Cet article décrit brièvement et clairement les trois lois de Mendel. Ces lois sont à la base de toute génétique ; en les créant, Mendel a réellement créé cette science.
Vous trouverez ici une définition de chaque loi et apprendrez un peu de nouveau sur la génétique et la biologie en général.
Avant de commencer à lire l'article, vous devez comprendre que le génotype est la totalité des gènes d'un organisme et que le phénotype est ses caractéristiques externes.
Qui est Mendel et qu’a-t-il fait ?
Gregor Johann Mendel est un célèbre biologiste autrichien, né en 1822 dans le village de Gincice. Il a bien étudié, mais sa famille avait des difficultés financières. Pour y faire face, Johann Mendel décida en 1943 de devenir moine dans un monastère tchèque de la ville de Brno et y reçut le nom de Gregor.

Gregor Johann Mendel (1822 - 1884)
Plus tard, il étudie la biologie à l’Université de Vienne, puis décide d’enseigner la physique et l’histoire naturelle à Brno. Parallèlement, le scientifique s’intéresse à la botanique. Il a mené des expériences sur le croisement de pois. Sur la base des résultats de ces expériences, le scientifique a dérivé trois lois de l'hérédité, qui font l'objet de cet article.
Publiées dans l'ouvrage « Expériences avec des hybrides végétaux » en 1866, ces lois n'ont pas reçu une large publicité et l'ouvrage a été rapidement oublié. On ne s’en souvient qu’après la mort de Mendel en 1884. Vous savez déjà combien de lois il a dérivées. Il est maintenant temps de passer à l'examen de chacun d'entre eux.
Première loi de Mendel - la loi d'uniformité des hybrides de première génération
Considérez l'expérience menée par Mendel. Il a pris deux types de pois. Ces espèces se distinguaient par la couleur de leurs fleurs. L’un les avait violets et l’autre blanc.
Après les avoir croisés, le scientifique a vu que tous les descendants avaient des fleurs violettes. Et les pois jaunes et verts ont produit une progéniture complètement jaune. Le biologiste a répété l’expérience plusieurs fois, vérifiant l’hérédité de différents traits, mais le résultat était toujours le même.

Sur la base de ces expériences, le scientifique a dérivé sa première loi, voici sa formulation : tous les hybrides de la première génération héritent toujours d'un seul trait de leurs parents.
Désignons par A le gène responsable des fleurs violettes et par a des fleurs blanches. Le génotype d'un parent est AA (violet) et le second est aa (blanc). Le gène A sera hérité du premier parent et a du second. Cela signifie que le génotype de la progéniture sera toujours Aa. Un gène désigné par une lettre majuscule est dit dominant, et une lettre minuscule est dit récessif.
Si le génotype d'un organisme contient deux gènes dominants ou deux gènes récessifs, alors il est dit homozygote, et un organisme contenant des gènes différents est appelé hétérozygote. Si l'organisme est hétérozygote, alors le gène récessif, appelé lettre capitale, est supprimé par un dominant plus fort, ce qui entraîne la manifestation du trait dont le dominant est responsable. Cela signifie que les pois de génotype Aa auront des fleurs violettes.
Le croisement de deux organismes hétérozygotes présentant des caractéristiques différentes est un croisement monohybride.
Codominance et dominance incomplète
Il arrive que gène dominant ne peut pas supprimer le récessif. Et puis les deux caractéristiques parentales apparaissent dans le corps.
Ce phénomène peut être observé dans l’exemple du camélia. Si dans le génotype de cette plante un gène est responsable des pétales rouges et l'autre du blanc, alors la moitié des pétales du camélia deviendront rouges et le reste blanc.

Ce phénomène est appelé codominance.
La domination incomplète est un phénomène similaire, dans lequel apparaît une troisième caractéristique, quelque chose entre ce qu'avaient les parents. Par exemple, une fleur de beauté nocturne avec un génotype contenant à la fois des pétales blancs et rouges devient rose.
La deuxième loi de Mendel - la loi de la ségrégation
Ainsi, nous nous souvenons que lors du croisement de deux organismes homozygotes, tous les descendants n'adopteront qu'un seul trait. Mais que se passerait-il si nous prenions deux organismes hétérozygotes de cette progéniture et les croisions ? La progéniture sera-t-elle uniforme ?

Revenons aux pois. Chaque parent est également susceptible de transmettre soit le gène A, soit le gène a. Ensuite, la descendance sera répartie comme suit :
- AA - fleurs violettes (25 %) ;
- aa - fleurs blanches (25%) ;
- Aa - fleurs violettes (50%).
On constate qu'il y a trois fois plus d'organismes à fleurs violettes. Il s'agit d'un phénomène de clivage. C'est la deuxième loi de Gregor Mendel : lors du croisement d'organismes hétérozygotes, la progéniture est divisée dans un rapport de 3:1 en phénotype et de 1:2:1 en génotype.
Il existe cependant des gènes dits mortels. S'ils sont présents, il y a un écart par rapport à la deuxième loi. Par exemple, la progéniture des souris jaunes est divisée dans un rapport de 2 : 1.
La même chose se produit avec les renards de couleur platine. Le fait est que si dans le génotype de ces organismes (et de certains autres), les deux gènes sont dominants, ils meurent tout simplement. En conséquence, un gène dominant ne peut être exprimé que si l’organisme est hétérozyote.
La loi de la pureté des gamètes et ses bases cytologiques
Prenons les pois jaunes et les pois verts, le gène jaune est dominant et le gène vert est récessif. L’hybride contiendra ces deux gènes (même si nous ne verrons que la manifestation du dominant).

On sait que les gènes sont transférés du parent à la progéniture à l'aide de gamètes. Un gamète est une cellule sexuelle. Il y a deux gènes dans le génotype hybride ; il s'avère que chaque gamète - et il y en a deux - contenait un gène. Après avoir fusionné, ils ont formé un génotype hybride.
Si, à la deuxième génération, un trait récessif caractéristique de l'un des organismes parents apparaissait, alors les conditions suivantes étaient remplies :
- les facteurs héréditaires des hybrides n'ont pas changé ;
- chaque gamète contenait un gène.
Le deuxième point est la loi de la pureté des gamètes. Bien sûr, il n’y a pas deux gènes, il y en a plusieurs. Il existe un concept de gènes alléliques. Ils sont responsables du même signe. Connaissant ce concept, nous pouvons formuler la loi comme suit : un gène choisi au hasard parmi un allèle pénètre dans le gamète.
La base cytologique de cette règle : les cellules dans lesquelles se trouvent des chromosomes contenant des paires d'allèles avec toute l'information génétique, se divisent et forment des cellules dans lesquelles il n'y a qu'un seul allèle - les cellules haploïdes. Dans ce cas, ce sont des gamètes.
La troisième loi de Mendel - la loi de l'héritage indépendant
Le respect de la troisième loi est possible avec le croisement dihybride, lorsque non pas un trait est étudié, mais plusieurs. Dans le cas des pois, il s’agit par exemple de la couleur et de la douceur des graines.

Nous désignons les gènes responsables de la couleur des graines par A (jaune) et a (vert) ; pour la douceur - B (lisse) et b (ridée). Essayons de réaliser un croisement dihybride d'organismes présentant des caractéristiques différentes.
La première loi n'est pas violée lors d'un tel croisement, c'est-à-dire que les hybrides seront identiques à la fois en termes de génotype (AaBb) et de phénotype (avec des graines jaunes et lisses).
Quelle sera la répartition de la deuxième génération ? Pour le savoir, vous devez découvrir quels gamètes les organismes parents peuvent sécréter. Ce sont évidemment AB, Ab, aB et ab. Après cela, un circuit appelé réseau de Pinnett est construit.
Tous les gamètes pouvant être libérés par un organisme sont répertoriés horizontalement, et tous les gamètes pouvant être libérés par un autre sont répertoriés verticalement. À l’intérieur de la grille est enregistré le génotype de l’organisme qui apparaîtrait avec les gamètes donnés.
| UN B | Un B | un B | un B | |
| UN B | AABB | AABb | AaBB | AaBb |
| Un B | AABb | AAbb | AaBb | Aabb |
| un B | AaBB | AaBb | aaBB | aaBb |
| un B | AaBb | Aabb | aaBb | aabb |
Si vous étudiez le tableau, vous pouvez conclure que la division des hybrides de deuxième génération par phénotype se produit dans le rapport 9:3:3:1. Mendel s'en est également rendu compte après avoir mené plusieurs expériences.
En outre, il est également arrivé à la conclusion que lequel des gènes d'un allèle (Aa) pénètre dans le gamète ne dépend pas de l'autre allèle (Bb), c'est-à-dire qu'il n'y a qu'un héritage indépendant de traits. C'est sa troisième loi, appelée loi de l'héritage indépendant.
Conclusion
Les trois lois de Mendel constituent les lois génétiques fondamentales. Grâce au fait qu'une personne a décidé d'expérimenter avec les pois, la biologie a reçu une nouvelle section - la génétique.
Grâce à son aide, les scientifiques du monde entier ont appris beaucoup de choses, de la prévention des maladies au génie génétique. La génétique est l’une des branches les plus intéressantes et prometteuses de la biologie.
Signe- toute caractéristique d'un organisme, toute qualité ou propriété par laquelle un organisme peut être distingué d'un autre.
Signes alternatifs- des variantes mutuellement exclusives d'un même trait (exemple : couleur jaune et verte des graines de pois).
Domination- la prédominance du trait d'un de ses parents chez un hybride.
Trait dominant- le trait prédominant qui apparaît dans la première génération de descendance chez les individus hétérozygotes et les homozygotes dominants (voir ci-dessous).
Trait récessif- un trait hérité, mais supprimé et n'apparaissant pas chez les descendants hétérozygotes ; se manifeste par l'état homozygote d'un gène récessif.
Phénotype- la totalité de tous les externes et signes internes corps. Le phénotype se forme grâce à l’interaction du génotype avec l’environnement de l’organisme.
Gènes alléliques, dominants et récessifs. Génotype
Allèle- l'une des formes alternatives d'existence d'un gène qui détermine un certain trait. Le nombre d'allèles d'un même gène peut atteindre plusieurs dizaines.
■ Chaque chromosome ou chromatide ne peut porter qu'un seul allèle d'un gène donné.
■ Seuls deux allèles de chaque gène sont présents dans les cellules d'un individu.
Lieu- la région du chromosome sur laquelle se situe le gène.
Gènes alléliques- des gènes situés dans les mêmes locus de chromosomes homologues et responsables de manifestations alternatives d'un même trait (exemple : gènes responsables de la couleur des yeux humains). Les gènes alléliques indiquent lettres identiques Alphabet latin : Un, un; B, b.
Gènes non alléliques- des gènes situés sur des chromosomes non homologues ou à différents locus de chromosomes homologues.
Gènes dominants- les gènes correspondant aux traits dominants ; sont désignés par des lettres majuscules latines (A, B).
Gènes récessifs- les gènes correspondant à des traits récessifs ; sont indiqués par des lettres latines minuscules ( un B).
Génotype- la totalité de tous les gènes d'un organisme donné.
Croisement
Croisement- obtenir une progéniture par union artificielle matériel génétique différents parents (différentes cellules) dans une cellule.
Dossier génétique du croisement :
■ Première ligne: lettre R.(parents), génotype du corps féminin, signe de croisement x, génotype du corps masculin ; sous les désignations de génotypes, les caractéristiques des organismes peuvent être indiquées ;
■ deuxième ligne: lettre g(gamètes) et (sous les désignations de génotypes, en cercles) les gamètes d'individus femelles et mâles ;
■ troisième ligne : lettre F k (descendants), génotypes des descendants (les caractéristiques des organismes peuvent être indiquées sous les désignations de génotypes) ; k - numéro de génération.
Homozygote- un zygote contenant le même les allèles d'un gène sont dominants ( AA, homozygote dominant) ou récessif ( aa, homozygote récessif).
■ Un individu homozygote produit un type de gamète et ne se divise pas lorsqu'il est croisé.
Hétérozygote - zygote, contenant deux allèles différents du même gène ( Ahh).
■ Un individu hétérozygote dans sa progéniture produit une ségrégation pour ce caractère. Produit plusieurs types de gamètes.
Règle (hypothèse) de pureté des gamètes. Puisque chaque chromosome ou chromatide ne peut porter qu'un seul allèle d'un gène donné, lorsque les chromosomes (lors de la première division de la méiose) ou les chromatides (lors de la deuxième division de la méiose) divergent, un seul des allèles de chaque paire allélique part avec eux dans les cellules haploïdes des gamètes.
C'est pourquoi: tout gamète d'un organisme ne porte qu'un seul allèle de chaque gène, c'est-à-dire Les allèles des gamètes ne se mélangent pas.
Conséquences de la règle de pureté des gamètes :
■ homozygote le corps ne produit qu'un seul type de gamète :
un organisme hétérozygote pour une paire de gènes forme deux types de gamètes (des deux chromosomes homologues du zygote en cours de méiose, un chromosome est avec le gène UN- pénètre dans un gamète, dans l'autre - avec le gène UN- dans un autre gamète) : ![]()
Hybridation- le processus de croisement de deux organismes de la même espèce (hybridation intraspécifique) ou d'espèces ou genres différents (hybridation à distance).
Hybride- un organisme obtenu en croisant des organismes génétiquement différents.
Croix monohybride- croisement d'organismes qui diffèrent les uns des autres par des variantes alternatives d'un seul trait (une paire d'allèles).
Croix d'analyse- croisement de l'organisme étudié avec un organisme possédant un génotype homozygote récessif (et ne produisant qu'un seul type de gamètes à allèles récessifs). Permet de déterminer le génotype de l'organisme étudié. Utilisé en sélection végétale et animale.
Croisement dihybride- croisement d'organismes qui diffèrent les uns des autres par des versions alternatives de deux traits (deux paires d'allèles).
Croisement polyhybride- croisement d'organismes qui diffèrent les uns des autres dans des versions alternatives de trois caractéristiques ou plus.
Héritage enchaîné- héritage conjoint de gènes localisés sur le même chromosome ; les gènes forment des groupes de liaison.
Fractionnement des fonctionnalités- un certain rapport se manifestant parmi la progéniture de la deuxième génération et des suivantes entre le nombre d'individus caractérisés par des caractéristiques alternatives des formes parentales d'origine.
■ Spécifique relations quantitatives entre les nombres d'individus portant les caractéristiques de chacune des formes parentales sont déterminés selon que les organismes parents sont homozygotes ou hétérozygotes pour ces caractéristiques.
La première loi de Mendel
La première loi de Mendel (loi d'uniformité des hybrides de première génération, ou règle de dominance ) décrit le croisement d'individus homozygotes : p Lors du croisement d'individus homozygotes (issus de lignées pures de la même espèce) qui diffèrent par l'un d'une paire de caractères alternatifs, les hybrides de première génération résultants sont uniformes à la fois en termes de phénotype et de génotype.

Conséquence: si la première génération est uniforme dans l'un des traits alternatifs des individus parents, alors ce trait est dominant , et les individus parents homozygote selon des caractéristiques alternatives.
La deuxième loi de Mendel
La deuxième loi de Mendel(loi de ségrégation) décrit un croisement mono-hybride d'individus hétérozygotes : lorsque des hybrides de la première génération (c'est-à-dire des individus hétérozygotes) différant par l'une d'une paire de caractéristiques alternatives sont croisés, dans la deuxième génération, une scission est observée dans un rapport de 3 : 1 pour le phénotype et de 1 : 2 : 1 pour le génotype. 
Répartition phénotypique : trois parties de descendants de la deuxième génération avec dominant signe et une partie - avec récessif .
Répartition du génotype : une partie de la descendance est homozygote dominante (AA), deux parties de la descendance sont hétérozygotes (Ah) et une partie est constituée d'homozygotes récessifs (aaa).
Corollaires de la deuxième loi de Mendel :
■ si la progéniture des individus parentaux donne une répartition phénotypique proche de 3 : 1, alors les individus parentaux d'origine pour ces allèles hétérozygote ;
■ test croisé : si la progéniture des parents présente une division phénotypique proche de 1 : 1, alors l'un des parents était hétérozygote et l'autre était homozygote et portait une paire d'allèles récessifs.
La troisième loi de Mendel
La troisième loi de Mendel (loi de l'héritage indépendant des caractères ) décrit un croisement dihybride d'individus : Lors du croisement d'organismes homozygotes qui diffèrent par deux ou plusieurs paires de traits, on observe dans la deuxième génération un héritage indépendant des gènes de différentes paires alléliques et de leurs traits correspondants.
Ceux. chaque paire de gènes alléliques (et leurs traits alternatifs correspondants) est héritée indépendamment les uns des autres ( une autre formulation de la 3ème loi de Mendel ).
❖ Détermination des génotypes possibles et des probabilités de leur apparition chez les individus de la deuxième génération : dans un premier temps, le génotype de la première génération et le type de ses gamètes Gf1 sont déterminés (voir tableau),

après quoi les génotypes des individus et les probabilités de leur apparition sont déterminés à l'aide de Grilles Punnett .
Grille de Punnett- un tableau à l'aide duquel le fractionnement des traits hérités indépendamment est représenté et analysé. Les gamètes femelles sont écrits horizontalement dans la rangée supérieure de cette grille, les gamètes mâles sont écrits verticalement dans la colonne de gauche et les génotypes des individus filles sont enregistrés aux intersections des rangées et des colonnes.
Exemple: traversée homozygote individu de pois caractérisé par deux dominant signes - couleur jaune et forme lisse des graines - avec homozygote pois individuel ayant deux alternatives récessif caractéristique - couleur verte et forme ridée des graines.

Puisque, selon la troisième loi de Mendel, le fractionnement pour chaque caractéristique se produit indépendamment : par couleur (dans la deuxième génération) dans un rapport de 3 : 1 (voir la deuxième loi de Mendel), par forme - également dans un rapport de 3 : 1, puis répartition selon le phénotype, c'est-à-dire par combinaison de caractéristiques, est observé dans le rapport (3 : 1) 2 = 9 : 3 : 3 : 1 (neuf parties sur 16 sont des graines jaunes et lisses, trois parties sont jaunes ridées, trois autres parties sont vertes lisses et une partie sont des graines vertes ridées).
Des données du réseau de Punnett, il résulte que lors du croisement dihybride d'individus homozygotes (notamment pois) chez des individus de la deuxième génération, neuf génotypes différents (classes génotypiques), qui se répartissent en quatre classes phénotypiques. Les descendants dominants sur deux traits (graines de pois lisses jaunes) possèdent l'un des génotypes suivants (la probabilité d'apparition d'un génotype donné est indiquée entre parenthèses) : AABB (1/16), AAVv (2/16), AaBB(2/16) ou AaVv(4/16); dominant selon le premier et récessif selon le deuxième caractère (graines jaunes ridées) - AAbb(1/16) ou Aaww(2/16); récessif selon le premier et dominant selon le deuxième caractère (graines vertes lisses) - aaBB(1/16) ou aaVv(2/16); récessif pour les deux caractères - génotype aabv (1/16) (graines ridées vertes).
❖ La ségrégation par génotype est la suivante :
■ quand dihybride
croisement : (1:2:1) 2 ;
■ quand polyhybride croisement (1:2:1) n, où n est le nombre de paires d'allèles en ségrégation.
❖ La division phénotypique ressemble à :
■ quand dihybride
croisement : (3 : 1) 2 = 9 : 3 : 1 ;
■ quand polyhybride
croisement (3 : 1) n.
Corollaires de la troisième loi de Mendel :
■ si l'analyse de la ségrégation pour deux caractères donne un rapport phénotypique proche de 9 : 3 : 3 : 1, alors les individus parentaux d'origine sont dihétérozygotes pour ces caractères ;
■ en général, chaque nouveau gène double le nombre de types de gamètes différents et triple le nombre de génotypes. Par conséquent, un individu hétérozygote pour n paires de gènes peut produire 2" types de gamètes et 3" génotypes différents ;
■ le nombre de classes différentes de phénotypes est égal au nombre de types différents de gamètes en présence de dominance et au nombre de génotypes différents en l'absence de dominance.
Remarques:
■ La troisième loi de Mendel, c'est-à-dire une combinaison indépendante de caractéristiques n'est réalisée qu'à la condition que les gènes alléliques qui déterminent ces caractéristiques soient situés dans différentes paires de chromosomes homologues ;
■ il n'explique pas les modes de transmission des gènes situés ensemble sur le même chromosome ;
❖ Calcul de la fréquence d'un génotype spécifique chez la progéniture de parents différant par un certain nombre de gènes hérités indépendamment :
■ Premièrement, la probabilité d'apparition du génotype correspondant est calculée séparément pour chaque paire de gènes ;
■ la fréquence souhaitée est égale au produit de ces probabilités. Exemple : calculer la fréquence du génotype Aabbcc dans la descendance du croisement AaBbcc x AaBbCc. La probabilité d'apparition du génotype Aa chez la descendance issue du croisement Aa x Aa est de 1/2 ; la probabilité d'apparition du génotype bb dans la descendance du croisement Bb x Bb est de 1/4 ; la probabilité d'apparition du génotype Cc dans la descendance d'un croisement Cc x CC est de 1/2. Par conséquent, la probabilité d’apparition du génotype AabbCC est de (1/2) x (1/4) x (1/2) = 1/16.
Conditions d'accomplissement et signification des lois de Mendel
Les lois de Mendel ne sont remplies qu'en moyenne, avec un grand nombre d'expériences du même type. Ils sont une conséquence de la combinaison aléatoire de gamètes porteurs de gènes différents, et de la nature statistique de l'hérédité, déterminée par un grand nombre de rencontres également probables de gamètes.
❖ Conditions supplémentaires, sous lequel les lois de Mendel sont satisfaites :
■ un gène doit contrôler un seul caractère, et un caractère doit être le résultat de l'action d'un seul gène ;
■ la domination doit être complète ;
■ il ne doit y avoir aucun lien entre les gènes ;
■ probabilité égale de formation de gamètes et de zygotes de types différents ;
■ probabilité égale de survie des descendants de génotypes différents ;
■ nombre de croisements statistiquement élevé.
❖ Le sens des lois de Mendel :
■ ces lois sont de nature universelle et ne dépendent pas de la position systématique de l'organisme et de la complexité de sa structure ;
■ avec leur aide, vous pouvez calculer le nombre de types de gamètes formés et établir des combinaisons possibles de traits dominants et récessifs chez les hybrides.
La génétique- une science qui étudie l'hérédité et la variabilité des organismes.
Hérédité- la capacité des organismes à transmettre leurs caractéristiques de génération en génération (caractéristiques de structure, fonction, développement).
Variabilité- la capacité des organismes à acquérir de nouvelles caractéristiques. L'hérédité et la variabilité sont deux propriétés opposées mais interdépendantes d'un organisme.
Hérédité
Concepts de base
Gène et allèles. L'unité d'information héréditaire est le gène.
Gène(du point de vue de la génétique) - une section d'un chromosome qui détermine le développement d'une ou plusieurs caractéristiques dans un organisme.
Allèles- différents états d'un même gène, situés dans un certain locus (région) de chromosomes homologues et déterminant le développement d'un trait particulier. Les chromosomes homologues ne sont présents que dans les cellules contenant un ensemble diploïde de chromosomes. On ne les trouve pas dans les cellules sexuelles (gamètes) des eucaryotes ou des procaryotes.
Panneau (sèche-cheveux)- une qualité ou une propriété par laquelle un organisme peut être distingué d'un autre.
Domination- le phénomène de prédominance du trait d'un des parents dans un hybride.
Trait dominant- un trait qui apparaît dès la première génération d'hybrides.
Trait récessif- un trait qui disparaît extérieurement dans la première génération d'hybrides.
Traits dominants et récessifs chez l'homme
| Panneaux | |
| dominant | récessif |
| Nanisme | Hauteur normale |
| Polydactylie (polydactylie) | Norme |
| Cheveux bouclés | Cheveux raides |
| Pas les cheveux roux | cheveux roux |
| Calvitie précoce | Norme |
| Long cils | Cils courts |
| Grands yeux | Petits yeux |
| yeux marrons | Yeux bleus ou gris |
| Myopie | Norme |
| Vision crépusculaire (cécité nocturne) | Norme |
| Taches de rousseur sur le visage | Pas de taches de rousseur |
| Coagulation sanguine normale | Mauvaise coagulation sanguine (hémophilie) |
| Vision des couleurs | Manque de vision des couleurs (daltonisme) |
Allèle dominant
- un allèle qui détermine un trait dominant. Indiqué par une lettre majuscule latine : A, B, C, ….
Allèle récessif
- un allèle qui détermine un trait récessif. Désigné par une lettre minuscule latine : a, b, c, ….
L'allèle dominant assure le développement du trait à la fois dans les états homozygotes et hétérozygotes, tandis que l'allèle récessif ne se manifeste qu'à l'état homozygote.
Homozygote et hétérozygote.
Les organismes (zygotes) peuvent être homozygotes ou hétérozygotes.
Organismes homozygotes
ont deux allèles identiques dans leur génotype - tous deux dominants ou tous deux récessifs (AA ou aa).
Organismes hétérozygotes
avoir l'un des allèles sous une forme dominante et l'autre sous une forme récessive (Aa).
Les individus homozygotes ne produisent pas de clivage dans la génération suivante, tandis que les individus hétérozygotes produisent un clivage.
Différentes formes alléliques de gènes résultent de mutations. Un gène peut muter à plusieurs reprises, produisant de nombreux allèles.
Allélisme multiple
- le phénomène d'existence de plus de deux formes alléliques alternatives d'un gène, ayant des manifestations différentes dans le phénotype. Deux ou plusieurs affections génétiques résultent de mutations. Une série de mutations provoque l'apparition d'une série d'allèles (A, a1, a2, ..., an, etc.), qui sont dans des relations dominantes-récessives différentes les unes par rapport aux autres.
Génotype
- la totalité de tous les gènes d'un organisme.
Phénotype
- l'ensemble de toutes les caractéristiques d'un organisme. Il s'agit notamment des caractéristiques morphologiques (externes) (couleur des yeux, couleur des fleurs), biochimiques (forme de la molécule protéine structurelle ou enzyme), histologiques (forme et taille des cellules), anatomiques, etc. En revanche, les signes peuvent être divisés en qualitatifs (couleur des yeux) et quantitatifs (poids corporel). Le phénotype dépend du génotype et des conditions environnementales. Il se développe à la suite de l’interaction du génotype et des conditions environnementales. Ces dernières influencent dans une moindre mesure les caractéristiques qualitatives et davantage les caractéristiques quantitatives.
Croisement (hybridation).
L'une des principales méthodes de génétique est le croisement ou l'hybridation.
Méthode hybridologique
- croisement (hybridation) d'organismes qui diffèrent les uns des autres par une ou plusieurs caractéristiques.
Hybrides
- les descendants issus de croisements d'organismes qui diffèrent les uns des autres par une ou plusieurs caractéristiques.
Selon le nombre de caractéristiques par lesquelles les parents diffèrent les uns des autres, ils distinguent différents types traversée.
Croix monohybride
- les croisements dans lesquels les parents ne diffèrent que par une seule caractéristique.
Croisement dihybride
- croisement dans lequel les parents diffèrent par deux caractéristiques.
Croisement polyhybride
- croisement dans lequel les parents diffèrent par plusieurs caractéristiques.
Pour enregistrer les résultats des croisements, les notations généralement acceptées suivantes sont utilisées :
R - parents (de lat. parental- parent);
F - progéniture (de lat. filial- progéniture) : F 1 - hybrides de première génération - descendants directs des parents P ; F 2 - hybrides de deuxième génération - descendants issus du croisement d'hybrides F 1 entre eux, etc.
♂ - mâle (bouclier et lance - signe de Mars) ;
♀ - femelle (miroir avec anse - signe de Vénus) ;
X - icône de croisement ;
: - le fractionnement des hybrides, sépare les rapports numériques des classes de descendance qui diffèrent (par phénotype ou génotype).
La méthode hybridologique a été développée par le naturaliste autrichien G. Mendel (1865). Il a utilisé des plants de pois de jardin autogames. Mendel a croisé des lignées pures (individus homozygotes) qui différaient les unes des autres par une, deux ou plusieurs caractéristiques. Il a obtenu des hybrides de première, deuxième génération, etc. Mendel a traité mathématiquement les données obtenues. Les résultats obtenus ont été formulés sous forme de lois de l'hérédité.
Les lois de G. Mendel
Première loi de Mendel. G. Mendel a croisé des plants de pois à graines jaunes et des plants de pois à graines vertes. Tous deux étaient des lignées pures, c’est-à-dire homozygotes.
Première loi de Mendel - la loi d'uniformité des hybrides de première génération (loi de dominance) : Lorsque des lignées pures sont croisées, tous les hybrides de première génération présentent un trait (dominant).
Deuxième loi de Mendel. Après cela, G. Mendel a croisé les hybrides de première génération entre eux.
La deuxième loi de Mendel est la loi du dédoublement des caractères : les hybrides de la première génération, lorsqu'ils sont croisés, sont divisés dans un certain rapport numérique : les individus présentant une manifestation récessive du trait représentent 1/4 des nombre total descendance.
La ségrégation est un phénomène dans lequel le croisement d'individus hétérozygotes conduit à la formation de descendants, dont certains portent un trait dominant et d'autres un trait récessif. Dans le cas d'un croisement monohybride, ce rapport est le suivant : 1AA:2Aa:1aa, soit 3:1 (en cas de dominance complète) ou 1:2:1 (en cas de dominance incomplète). Dans le cas d'un croisement dihybride - 9:3:3:1 ou (3:1) 2. Avec polyhybride - (3:1) n.
Dominance incomplète. Un gène dominant ne supprime pas toujours complètement un gène récessif. Ce phénomène est appelé dominance incomplète
. Un exemple de domination incomplète est l'héritage de la couleur des fleurs de beauté nocturne.
Bases cytologiques de l'uniformité de la première génération et du clivage des caractères de la deuxième génération consistent en la divergence de chromosomes homologues et la formation de cellules germinales haploïdes lors de la méiose.
Hypothèse (loi) de pureté des gamètes déclare : 1) lors de la formation des cellules germinales, un seul allèle d'une paire allélique pénètre dans chaque gamète, c'est-à-dire que les gamètes sont génétiquement purs ; 2) dans un organisme hybride, les gènes ne s'hybrident pas (ne se mélangent pas) et sont dans un état allélique pur.
Caractère statistique des phénomènes de clivage. De l'hypothèse de la pureté des gamètes, il résulte que la loi de ségrégation est le résultat d'une combinaison aléatoire de gamètes porteurs de gènes différents. Compte tenu du caractère aléatoire de la connexion des gamètes, le résultat global s'avère naturel. Il s'ensuit que dans le croisement monohybride, le rapport de 3:1 (dans le cas d'une dominance complète) ou de 1:2:1 (dans le cas d'une dominance incomplète) doit être considéré comme un modèle basé sur des phénomènes statistiques. Ceci s'applique également au cas de croisement polyhybride. La mise en œuvre précise des relations numériques lors de la division n'est possible qu'avec un grand nombre d'individus hybrides étudiés. Ainsi, les lois de la génétique sont de nature statistique.
Analyse de la progéniture. Croix d'analyse permet de déterminer si un organisme est homozygote ou hétérozygote pour un gène dominant. Pour ce faire, un individu dont il faut déterminer le génotype est croisé avec un individu homozygote pour le gène récessif. Souvent, l'un des parents est croisé avec l'un des descendants. Cette traversée s'appelle consigné
.
En cas d'homozygotie de l'individu dominant, le clivage n'aura pas lieu :
En cas d'hétérozygotie de l'individu dominant, un clivage se produira :
Troisième loi de Mendel. G. Mendel a réalisé un croisement dihybride de plants de pois à graines jaunes et lisses et de plants de pois à graines vertes et ridées (les deux sont des lignées pures), puis a croisé leurs descendants. En conséquence, il a découvert que chaque paire de traits, lorsqu’elle est divisée dans la progéniture, se comporte de la même manière que dans un croisement monohybride (séparation 3:1), c’est-à-dire indépendamment de l’autre paire de traits.
La troisième loi de Mendel- la loi de la combinaison indépendante (héritage) des traits : le dédoublement pour chaque trait se produit indépendamment des autres traits.
Base cytologique de combinaison indépendante est le caractère aléatoire de la divergence des chromosomes homologues de chaque paire vers différents pôles de la cellule au cours du processus de méiose, indépendamment des autres paires de chromosomes homologues. Cette loi n'est valable que si les gènes responsables du développement de différents traits sont situés sur des chromosomes différents. Les exceptions sont les cas d'héritage lié.
Héritage enchaîné. Perte d'adhérence
Le développement de la génétique a montré que tous les traits ne sont pas hérités conformément aux lois de Mendel. Ainsi, la loi de l'hérédité indépendante des gènes n'est valable que pour les gènes situés sur des chromosomes différents.
Les modèles de transmission génétique liée ont été étudiés par T. Morgan et ses étudiants au début des années 20. XXe siècle L'objet de leurs recherches était la mouche des fruits, la drosophile (sa durée de vie est courte et plusieurs dizaines de générations peuvent être obtenues en un an ; son caryotype est constitué de seulement quatre paires de chromosomes).
La loi de Morgan : les gènes localisés sur le même chromosome sont hérités principalement ensemble.
Gènes liés
- des gènes situés sur le même chromosome.
Groupe d'embrayage
- tous les gènes sur un chromosome.
Dans un certain pourcentage de cas, l'adhésion peut être rompue. La raison de la rupture de la cohésion est le croisement (croisement des chromosomes) - l'échange de sections chromosomiques dans la prophase I de la division méiotique. La traversée mène à Recombinaison génétique. Plus les gènes sont éloignés les uns des autres, plus les croisements se produisent souvent entre eux. La construction est basée sur ce phénomène cartes génétiques- détermination de la séquence des gènes sur le chromosome et de la distance approximative qui les sépare.
Génétique du sexe
Autosomes
- des chromosomes identiques chez les deux sexes.
Chromosomes sexuels (hétérochromosomes)
- les chromosomes sur lesquels les sexes masculin et féminin diffèrent.
Une cellule humaine contient 46 chromosomes, soit 23 paires : 22 paires d'autosomes et 1 paire de chromosomes sexuels. Les chromosomes sexuels sont appelés chromosomes X et Y. Les femmes ont deux chromosomes X et les hommes un chromosome X et un chromosome Y.
Il existe 5 types de détermination du sexe chromosomique.
Types de détermination du sexe chromosomique
| Taper | Exemples |
| ♀ XX, ♂ ХY | Caractéristiques des mammifères (y compris les humains), des vers, des crustacés, de la plupart des insectes (y compris les mouches des fruits), de la plupart des amphibiens et de certains poissons. |
| ♀XY, ♂XX | Caractéristique des oiseaux, reptiles, certains amphibiens et poissons, certains insectes (Lépidoptères) |
| ♀ XX, ♂ X0 | Présent chez certains insectes (orthoptères); 0 signifie pas de chromosomes |
| ♀ X0, ♂ XX | Trouvé chez certains insectes (homoptères) |
| type haplo-diploïde (♀ 2n, ♂ n) | On le retrouve par exemple chez les abeilles et les fourmis : les mâles se développent à partir d'œufs haploïdes non fécondés (parthénogenèse), les femelles à partir d'œufs diploïdes fécondés. |
Héritage lié au sexe
- héritage de traits dont les gènes sont situés sur les chromosomes X et Y. Les chromosomes sexuels peuvent contenir des gènes qui ne sont pas liés au développement des caractéristiques sexuelles.
Dans une combinaison XY, la plupart des gènes présents sur le chromosome X n'ont pas de paire allélique sur le chromosome Y. De plus, les gènes situés sur le chromosome Y n’ont pas d’allèles sur le chromosome X. De tels organismes sont appelés hémizygote
. Dans ce cas, un gène récessif apparaît, présent au singulier dans le génotype. Ainsi, le chromosome X peut contenir un gène responsable de l’hémophilie (diminution de la coagulation sanguine). Ensuite, tous les hommes ayant reçu ce chromosome souffriront de cette maladie, puisque le chromosome Y ne contient pas d'allèle dominant.
Génétique du sang
Selon le système ABO, les gens ont 4 groupes sanguins. Le groupe sanguin est déterminé par le gène I. Chez l'homme, le groupe sanguin est déterminé par trois gènes IA, IB, I0. Les deux premiers sont codominants l’un par rapport à l’autre et tous deux sont dominants par rapport au troisième. En conséquence, une personne possède 6 groupes sanguins selon la génétique et 4 selon la physiologie.
| Groupe I | 0 | je 0 je 0 | homozygote |
| Groupe II | UN | je a je a | homozygote |
| je un je 0 | hétérozygote | ||
| Groupe III | DANS | Je B Je B | homozygote |
| Je B Je 0 | hétérozygote | ||
| Groupe IV | UN B | Je A I B | hétérozygote |
U différentes nations Le rapport des groupes sanguins dans la population est différent.
Répartition des groupes sanguins selon le système AB0 dans différents pays,%
De plus, le sang personnes différentes peut différer selon le facteur Rh. Le sang peut être Rh positif (Rh +) ou Rh négatif (Rh -). Ce ratio varie selon les nations.
Répartition du facteur Rh parmi différents peuples,%
| Nationalité | Rh positif | Rh négatif |
| Aborigènes australiens | 100 | 0 |
| Indiens d'Amérique | 90–98 | 2–10 |
| Arabes | 72 | 28 |
| Basque | 64 | 36 |
| Chinois | 98–100 | 0–2 |
| Mexicains | 100 | 0 |
| norrois | 85 | 15 |
| les Russes | 86 | 14 |
| Esquimaux | 99–100 | 0–1 |
| Japonais | 99–100 | 0–1 |
Le facteur Rh du sang est déterminé par le gène R. R + donne des informations sur la production de protéines (protéine Rh-positive), mais pas le gène R. Le premier gène est dominant sur le second. Si du sang Rh + est transfusé à une personne ayant du sang Rh –, des agglutinines spécifiques se forment chez elle et l'administration répétée de ce sang provoquera une agglutination. Lorsqu'une femme Rh développe un fœtus qui a hérité du Rh positif du père, un conflit Rh peut survenir. En règle générale, la première grossesse se termine en toute sécurité et la seconde se termine par une maladie de l'enfant ou une mortinatalité.
Interaction des gènes
Un génotype n’est pas simplement un ensemble mécanique de gènes. Il s’agit d’un système historiquement établi de gènes interagissant les uns avec les autres. Plus précisément, ce ne sont pas les gènes eux-mêmes (sections de molécules d'ADN) qui interagissent, mais les produits qui en sont formés (ARN et protéines).
Les gènes alléliques et non alléliques peuvent interagir.
Interaction des gènes alléliques :
dominance complète, dominance incomplète, codominance.
Domination totale
- un phénomène où un gène dominant supprime complètement le travail d'un gène récessif, entraînant le développement d'un trait dominant.
Dominance incomplète
- un phénomène lorsqu'un gène dominant ne supprime pas complètement le travail d'un gène récessif, à la suite duquel un trait intermédiaire se développe.
Codominance
(manifestation indépendante) est un phénomène lorsque les deux allèles participent à la formation d'un trait dans un organisme hétérozygote. Chez l’homme, le gène qui détermine le groupe sanguin est représenté par une série d’allèles multiples. Dans ce cas, les gènes qui déterminent les groupes sanguins A et B sont codominants l'un par rapport à l'autre, et tous deux sont dominants par rapport au gène qui détermine le groupe sanguin 0.
Interaction des gènes non alléliques :
coopération, complémentarité, épistase et polymère.
Coopération
- un phénomène où, en raison de l'action mutuelle de deux gènes dominants non alléliques, dont chacun a sa propre manifestation phénotypique, un nouveau trait se forme.
Complémentarité
- un phénomène lorsqu'un trait se développe uniquement par l'action mutuelle de deux gènes dominants non alléliques, dont chacun ne provoque pas individuellement le développement du trait.
Épistase
- un phénomène lorsqu'un gène (à la fois dominant et récessif) supprime l'action d'un autre gène (non allélique) (à la fois dominant et récessif). Le gène suppresseur peut être dominant (épistasie dominante) ou récessif (épistasie récessive).
Polymérisme
- un phénomène où plusieurs gènes dominants non alléliques sont responsables d'effets similaires sur le développement d'un même trait. Plus ces gènes sont présents dans le génotype, plus le caractère est prononcé. Le phénomène de polymérisation s'observe lors de la transmission de caractères quantitatifs (couleur de peau, poids corporel, rendement laitier des vaches).
Contrairement à la polymérisation, il existe un phénomène tel que pléiotropie
- l'action de plusieurs gènes, lorsqu'un gène est responsable du développement de plusieurs caractères.
Théorie chromosomique de l'hérédité
Dispositions de base de la théorie chromosomique de l'hérédité :
- Les chromosomes jouent un rôle de premier plan dans l'hérédité ;
- les gènes sont situés sur le chromosome dans une certaine séquence linéaire ;
- chaque gène est situé à un endroit précis (locus) du chromosome ; les gènes alléliques occupent des locus identiques sur des chromosomes homologues ;
- les gènes des chromosomes homologues forment un groupe de liaison ; leur nombre est égal à l'ensemble haploïde des chromosomes ;
- l'échange de gènes alléliques (crossing over) est possible entre chromosomes homologues ;
- La fréquence de croisement entre les gènes est proportionnelle à la distance qui les sépare.
Héritage non chromosomique
Selon la théorie chromosomique de l’hérédité, l’ADN des chromosomes joue un rôle majeur dans l’hérédité. Cependant, l'ADN est également contenu dans les mitochondries, les chloroplastes et le cytoplasme. L'ADN non chromosomique est appelé plasmides . Les cellules ne disposent pas de mécanismes spéciaux pour une distribution uniforme des plasmides lors de la division, de sorte qu'une cellule fille peut recevoir une information génétique et la seconde - complètement différente. L'hérédité des gènes contenus dans les plasmides n'obéit pas aux lois mendéliennes de l'hérédité, et leur rôle dans la formation du génotype n'a pas encore été suffisamment étudié.
Modèles d'hérédité. Les lois de G. Mendel, leur nature statistique et leurs fondements cytologiques
Les lois fondamentales de l'hérédité ont été établies par l'éminent scientifique tchèque Gregor Mendel. G. Mendel a commencé ses recherches avec le croisement monohybride, dans lequel les individus parents diffèrent par l'état d'un trait. Le pois de semence qu'il a choisi est une plante auto-reproductrice, les descendants de chaque individu sont donc des lignées pures. Ensemble, les pois peuvent être artificiellement pollinisés, ce qui permet de s'hybrider et d'obtenir des formes hétérozygotes (hybrides). Les plantes d'une lignée pure avec des graines de couleur jaune ont été considérées comme des plantes maternelles (P) et parentales (P) de couleur verte. À la suite d'un tel croisement, les graines des plantes (hybrides de première génération - F1) se sont révélées uniformes - de couleur jaune. Autrement dit, seuls les traits dominants sont apparus dans le phénotype des hybrides F1.
L’uniformité de la première génération hybride et l’identification d’un seul trait dominant chez les hybrides est appelée loi de dominance ou loi de Mendel.
La ségrégation est le phénomène de manifestation des deux états de traits dans la deuxième génération d'hybrides (F2), en raison de la différence dans les gènes alléliques qui les déterminent.
Il existe des plantes F1 autogames à graines jaunes qui produisent une progéniture avec des graines jaunes et vertes ; le trait récessif ne disparaît pas, mais n'est que temporairement supprimé et réapparaît en F2 dans la proportion de 1/4 de graines vertes et 3/4 de graines jaunes. C'est exactement - 3:1.
La manifestation d’un trait récessif dans le phénotype d’un quart des hybrides de deuxième génération, et la manifestation d’un trait dominant chez les trois quarts, est appelée loi de ségrégation, loi de Mendel II.
Par la suite, G. Mendel a compliqué les conditions des expériences - il a utilisé des plantes qui différaient par différents états de deux (croisement dihybride) ou plus traits (croisement polyhybride). Lors du croisement de plants de pois avec des graines jaunes lisses et des graines vertes ridées, tous les hybrides de première génération avaient des graines jaunes lisses - une manifestation de la loi de Mendel - l'uniformité des hybrides de première génération. Mais parmi les hybrides F2, il y avait quatre phénotypes.
Sur la base des résultats obtenus, G. Mendel a formulé la loi de combinaison indépendante des états de caractère (la loi de l'héritage indépendant des caractères). C'est la troisième loi de Mendel. Dans le croisement di- ou polyhybride, la division des états de chaque trait dans la descendance se produit indépendamment des autres. Les croisements dihybrides sont caractérisés par un clivage selon le phénotype 9:3:3:1, et des groupes avec de nouvelles combinaisons de caractéristiques apparaissent.
La domination incomplète est la nature intermédiaire de l'héritage. Il existe des allèles qui ne sont que partiellement dominants sur les allèles récessifs. L’individu hybride possède alors un certain degré de trait dans le phénotype, qui le distingue de ses parents. Ce phénomène est appelé dominance incomplète.
Méthodes de vérification du génotype des individus hybrides
Comme on le sait, avec une dominance complète, les individus possédant un ensemble de chromosomes dominant et hétérozygote sont phénotypiquement identiques. Il est possible de déterminer leur génotype par croisement analytique. Elle repose sur le fait que les individus homozygotes pour un trait récessif sont toujours similaires sur le plan phénotypique. Il s'agit du croisement d'un individu homozygote récessif avec un individu présentant un trait dominant mais un génotype inconnu.
Lors de la réception d’un F1 monotone, chaque parent ne produit qu’un seul type de gamète. Ainsi, l'individu dominant est homozygote pour le génotype (AA).
Si, lors du croisement d’un individu ayant un trait dominant avec un individu ayant un trait homozygote récessif, la progéniture résultante présente une division 1: 1, alors l’individu ayant le trait dominant est hétérozygote (Aa).
- Caractéristiques de la méthode d'analyse hybridologique. Les lois de Mendel.
- Types d'interactions génétiques.
- Héritage lié des traits.
- Héritage cytoplasmique.
Méthode analyse hybridologique , qui consiste à croiser et à comptabiliser ultérieurement les divisions (rapports de variétés phénotypiques et génotypiques de descendance), a été développé par le naturaliste tchèque G. Mendel (1865). Les caractéristiques de cette méthode comprennent : 1) lors du croisement, la prise en compte non pas de l'ensemble diversifié des traits chez les parents et les descendants, mais l'analyse de l'héritage des traits alternatifs individuels identifiés par le chercheur ; 2) comptabilité quantitative dans une série de générations successives de plantes hybrides qui diffèrent par leurs caractéristiques individuelles ; 3) analyse individuelle de la progéniture de chaque plante.
Travaillant avec des plants de pois de jardin autogames, G. Mendel a choisi pour l'expérimentation des variétés (lignées pures) qui différaient les unes des autres par des manifestations alternatives de caractères. Mendel a traité mathématiquement les données obtenues, ce qui a révélé un modèle clair d'héritage des caractéristiques individuelles des formes parentales par leurs descendants dans un certain nombre de générations suivantes. Mendel a formulé ce modèle sous la forme de règles d'hérédité, appelées plus tard Les lois de Mendel.
Le croisement de deux organismes s'appelle hybridation. Monohybride (monogène) est un croisement de deux organismes dans lequel l'héritage d'une paire de manifestations alternatives d'un trait est retracé (le développement de ce trait est déterminé par une paire d'allèles du même gène). Les hybrides de première génération sont uniformes dans le caractère étudié. En F1, une seule variante parmi une paire de variantes alternatives du caractère de couleur des graines apparaît, appelée dominant. Ces résultats illustrent la première loi de Mendel – la loi d'uniformité des hybrides de première génération, ainsi que la règle de dominance.
La première loi de Mendel peut être formulé comme suit : lors du croisement d'individus homozygotes qui diffèrent par une ou plusieurs paires de traits alternatifs, tous les hybrides de première génération seront uniformes dans ces traits. Les hybrides présenteront les traits dominants de leurs parents.
Dans la deuxième génération, une division selon le trait étudié a été découverte
Le ratio de descendance présentant des manifestations dominantes et récessives du trait s'est avéré être proche de ¾ à ¼. Ainsi, La deuxième loi de Mendel peut être formulé comme suit : lors d'un croisement monohybride d'individus hétérozygotes (hybrides F1) de deuxième génération, un clivage est observé dans les variantes du trait analysé dans un rapport de 3 :1 par phénotype et 1 :2 :1 par génotype. Pour expliquer la répartition des traits chez les hybrides des générations successives, G. Mendel a suggéré que chaque trait héréditaire dépend de la présence dans les cellules somatiques de deux facteurs héréditaires reçus du père et de la mère. Il est désormais établi que les facteurs héréditaires mendéliens correspondent à des gènes – les loci chromosomiques.
Les plantes homozygotes à graines jaunes (AA) produisent des gamètes de la même variété avec l'allèle A ; les plantes à graines vertes (aa) produisent des gamètes avec a. Ainsi, en utilisant la terminologie moderne, l’hypothèse « pureté des gamètes« peut être formulé comme suit : « Dans le processus de formation des cellules germinales, un seul gène d'une paire allélique pénètre dans chaque gamète, car pendant le processus de méiose, un chromosome d'une paire de chromosomes homologues pénètre dans le gamète.
Le croisement dans lequel l'héritage de deux paires de caractères alternatifs est tracé est appelé dihybride, selon plusieurs paires de signes - polyhybride. Dans les expériences de Mendel, lors du croisement d'une variété de pois à graines jaunes (A) et lisses (B) avec une variété de pois à graines vertes (a) et ridées (b), les hybrides F1 avaient des graines jaunes et lisses, c'est-à-dire Des caractères dominants apparaissent (les hybrides sont uniformes).
Les graines hybrides de la deuxième génération (F2) ont été réparties en quatre groupes phénotypiques dans le rapport : 315 - avec des graines jaunes lisses, 101 - avec des graines jaunes ridées, 108 - avec des graines vertes lisses, 32 - avec des graines vertes ridées. Si le nombre de descendants dans chaque groupe est divisé par le nombre de descendants dans le plus petit groupe, alors dans F2, le rapport des classes phénotypiques est d'environ 9:3:3:1. Ainsi, selon La troisième loi de Mendel, les gènes de différentes paires alléliques et les caractéristiques qui leur correspondent sont transmis à la progéniture indépendamment de de chacun d'eux combinant dans toutes sortes de combinaisons.
Avec une dominance complète d'un allèle sur l'autre, les individus hétérozygotes sont phénotypiquement impossibles à distinguer des individus homozygotes pour l'allèle dominant et ne peuvent être distingués que par une analyse hybridologique, c'est-à-dire selon la descendance issue d'un certain type de croisement, appelé en cours d'analyse. L'analyse est un type de croisement dans lequel un individu test présentant un trait dominant est croisé avec un individu homozygote pour la pomme récessive.
Si l'individu dominant est homozygote, la progéniture issue d'un tel croisement sera uniforme et aucune ségrégation ne se produira. Dans le cas où un individu présentant un trait dominant est hétérozygote, une division se produira dans un rapport de 1 : 1 en termes de phénotype et de génotype.
Interaction des gènes
Dans certains cas, l'action de différents gènes est relativement indépendante, mais, en règle générale, la manifestation de traits est le résultat de l'interaction de produits de différents gènes. Ces interactions peuvent être liées à la fois allélique, donc avec non allélique gènes.
Interaction entre allèles les gènes sont réalisés sous trois formes : dominance complète, dominance incomplète et manifestation indépendante (co-dominance).
Auparavant, les expériences de Mendel ont été examinées, qui ont révélé la dominance complète d'un allèle et la récessivité de l'autre. Une dominance incomplète est observée lorsqu'un gène d'une paire d'allèles ne permet pas la formation de son produit protéique suffisant pour la manifestation normale du trait. Avec cette forme d'interaction génétique, tous les hétérozygotes et homozygotes diffèrent considérablement les uns des autres par leur phénotype. À co-dominance dans les organismes hétérozygotes, chacun des gènes alléliques provoque la formation d'un trait contrôlé par lui dans le phénotype. Un exemple de cette forme d'interaction d'allèles est l'héritage des groupes sanguins humains selon le système ABO, déterminé par le gène I. Il existe trois allèles de ce gène, Io, Ia, Ib, qui déterminent les antigènes des groupes sanguins. La transmission des groupes sanguins illustre également le phénomène pluriel allélisme : dans le pool génétique des populations humaines, le gène I existe sous la forme de trois allèles différents, qui ne sont combinés chez les individus que par paires.
Interaction des gènes non alléliques. Dans certains cas, un trait d’un organisme peut être influencé par deux (ou plus) paires de gènes non alléliques. Cela conduit à des écarts numériques significatifs des classes phénotypiques (mais pas génotypiques) par rapport à celles établies par Mendel lors du croisement dihybride. L'interaction des gènes non alléliques se divise en formes principales : complémentarité, épistasie, polymérisation.
À complémentaire interaction, le trait ne se manifeste que dans le cas de la présence simultanée de deux gènes non alléliques dominants dans le génotype de l'organisme. Un exemple d’interaction complémentaire est le croisement de deux variétés différentes de pois de senteur aux pétales de fleurs blanches.
Le prochain type d'interaction de gènes non alléliques est l'épistasie, dans laquelle le gène d'une paire allélique supprime l'effet du gène de l'autre paire. Un gène qui supprime l'action d'un autre est appelé génome épistatique(ou suppresseur). Le gène supprimé est appelé hypostatique. L'épistase peut être dominante ou récessive. Un exemple d’épistasie dominante est l’héritage de la couleur du plumage chez les poulets. Le gène C sous sa forme dominante détermine la production normale de pigment, mais l'allèle dominant d'un autre gène I est son suppresseur. En conséquence, les poulets qui ont un allèle dominant du gène de couleur dans leur génotype se révèlent blancs en présence d'un suppresseur. L'effet épistatique d'un gène récessif illustre l'hérédité de la couleur du pelage chez la souris domestique. La couleur de l'agouti (couleur du pelage gris rougeâtre) est déterminée par le gène dominant A. Son allèle récessif a, à l'état homozygote, provoque une coloration noire. Le gène dominant d'une autre paire C détermine le développement du pigment ; les homozygotes pour l'allèle récessif c sont des albinos à fourrure blanche et aux yeux rouges (manque de pigment dans la fourrure et l'iris des yeux).
L'héritage d'un trait dont la transmission et le développement sont déterminés, en règle générale, par deux allèles d'un gène, est appelé monogénique. De plus, des gènes provenant de différentes paires alléliques sont connus (ils sont appelés polymères ou polygènes), ayant à peu près le même effet sur le trait.
Le phénomène d'action simultanée sur un trait de plusieurs gènes non alléliques du même type est appelé polymérisation. Bien que les gènes polymères ne soient pas alléliques, puisqu'ils déterminent le développement d'un trait, ils sont généralement désignés par une lettre A (a), avec des chiffres indiquant le nombre de paires alléliques. L'action des polygènes est le plus souvent additive.
Héritage enchaîné
Une analyse de la transmission simultanée de plusieurs traits chez la drosophile, réalisée par T. Morgan, a montré que les résultats des croisements analytiques d'hybrides F1 diffèrent parfois de ceux attendus dans le cas de leur transmission indépendante. Chez les descendants d'un tel croisement, au lieu de combiner librement les caractéristiques de différents couples, on a observé une tendance à hériter de combinaisons de caractéristiques principalement parentales. Cet héritage de traits était appelé lié. L'hérédité liée s'explique par la localisation des gènes correspondants sur le même chromosome. Dans le cadre de ces derniers, ils sont transmis de génération en génération de cellules et d'organismes, préservant la combinaison d'allèles des parents.
La dépendance de l'héritage lié des traits à la localisation des gènes sur un chromosome donne des raisons de considérer les chromosomes comme distincts groupes d'embrayage. Une analyse de l'héritage du trait de couleur des yeux chez la drosophile dans le laboratoire de T. Morgan a révélé certaines caractéristiques qui nous ont obligés à le distinguer comme un type distinct d'héritage de traits. héritage lié au sexe.
La dépendance des résultats expérimentaux sur le parent porteur de la variante dominante du trait nous a permis de suggérer que le gène qui détermine la couleur des yeux chez la drosophile est situé sur le chromosome X et n'a pas d'homologue sur le chromosome Y. Toutes les caractéristiques de l'héritage lié au sexe s'expliquent par la dose inégale des gènes correspondants chez les représentants de sexes différents - homo- et hétérogamétiques. Le chromosome X est présent dans le caryotype de chaque individu, c'est pourquoi les caractéristiques déterminées par les gènes de ce chromosome se forment chez les représentants féminins et masculins. Les individus du sexe homogamétique reçoivent ces gènes des deux parents et les transmettent à tous leurs descendants par l'intermédiaire de leurs gamètes. Les représentants du sexe hétérogamétique reçoivent un seul chromosome X du parent homogamétique et le transmettent à leur progéniture homogamétique. Chez les mammifères (y compris les humains), le sexe masculin reçoit de la mère les gènes liés à l'X et les transmet à ses filles. Dans le même temps, le sexe masculin n’hérite jamais du trait paternel lié à l’X et ne le transmet pas à ses fils.
Les gènes fonctionnant activement sur le chromosome Y, qui n'ont pas d'allèles sur le chromosome X, sont présents dans le génotype uniquement du sexe hétérogamétique et à l'état hémizygote. Par conséquent, ils se manifestent de manière phénotypique et ne se transmettent de génération en génération que chez les représentants du sexe hétérogamétique. Ainsi, chez l'homme, le signe de l'hypertrichose du pavillon d'oreille (« oreilles velues ») s'observe exclusivement chez l'homme et est hérité de père en fils.
Nous commencerons par présenter les lois de Mendel, puis nous parlerons de Morgan, et à la fin nous dirons pourquoi la génétique est nécessaire aujourd'hui, comment elle aide et quelles sont ses méthodes.
Dans les années 1860, le moine Mendel commença des recherches sur l’héritage des traits. Cela a été fait avant lui, et c’est la première fois que cela est mentionné dans la Bible. L'Ancien Testament dit que si un éleveur voulait acquérir une certaine race, il nourrissait certains moutons avec des branches pelées s'il voulait avoir une progéniture avec de la laine blanche, et des moutons non écossés s'il voulait avoir des peaux de bétail noires. Autrement dit, la façon dont les traits sont hérités inquiétait les gens avant même la rédaction de la Bible. Pourquoi, avant Mendel, n’ont-ils pas pu trouver des lois pour la transmission des traits à travers les générations ?
Le fait est qu'avant lui, les chercheurs ont choisi un ensemble de caractéristiques d'un individu, plus difficiles à gérer qu'un seul trait. Avant lui, la transmission des caractéristiques était souvent considérée comme un complexe unique (par exemple, elle a le visage d'une grand-mère, bien qu'il existe de nombreux signes individuels). Et Mendel a enregistré la transmission de chaque trait séparément, quelle que soit la manière dont les autres traits ont été transmis aux descendants.
Il est important que Mendel ait choisi pour l'étude des caractéristiques dont l'enregistrement était extrêmement simple. Ce sont des signes discrets et alternatifs :
- signes discrets (discontinus) : un signe donné est soit présent, soit absent. Par exemple, un attribut de couleur : un pois est vert ou pas vert.
- caractéristiques alternatives : un état d’un trait exclut la présence d’un autre état. Par exemple, l'état d'une caractéristique telle que la couleur : un pois est soit vert, soit jaune. Les deux états d’un trait ne peuvent pas apparaître dans un seul organisme.
Mendel avait une approche d'analyse des descendants qui n'avait pas été utilisée auparavant. Il s'agit d'une méthode d'analyse quantitative et statistique : tous les descendants avec un état de trait donné (par exemple, les pois verts) ont été regroupés en un seul groupe et leur nombre a été calculé, qui a été comparé au nombre de descendants avec un état de trait différent (jaune). petits pois).
Comme trait, Mendel a choisi la couleur des graines de pois, dont l'état s'excluait mutuellement : la couleur était soit jaune, soit verte. Un autre signe est la forme des graines. Les états alternatifs du trait sont que la forme est soit ridée, soit lisse. Il s’est avéré que ces caractéristiques se reproduisent de manière stable au fil des générations et apparaissent soit dans un état, soit dans un autre. Au total, Mendel a étudié 7 paires de traits, en surveillant chacun séparément.
En croisement, Mendel a étudié la transmission des traits des parents à leur progéniture. Et c'est ce qu'il a obtenu. Au cours de l'autopollinisation, l'un des parents n'a produit que des graines ridées au cours d'une série de générations, l'autre parent - uniquement des graines lisses.
Les pois sont autogames. Afin d’avoir une progéniture de deux parents différents (hybrides), il devait s’assurer que les plantes ne s’autopollinisent pas. Pour ce faire, il a retiré les étamines d’une plante mère et y a transféré le pollen d’une autre plante. Dans ce cas, les graines obtenues étaient hybrides. Toutes les graines hybrides de la première génération se sont révélées identiques. Ils se sont tous avérés lisses. Nous appelons l'état manifesté du trait dominant (le sens de la racine de ce mot est dominant). Un autre état du caractère (graines ridées) n’a pas été détecté chez les hybrides. Nous appelons cet état du trait récessif (inférieur).
Mendel a croisé en lui la première génération de plantes et a examiné la forme des pois résultants (c'était la deuxième génération de la progéniture du croisement). La majeure partie des graines s'est avérée lisse. Mais une partie était ridée, exactement la même que celle du parent d'origine (si nous parlions de notre propre famille, nous dirions que le petit-fils était exactement comme son grand-père, même si son père et sa mère n'avaient pas du tout ce signe). ). Il a mené une étude quantitative de la proportion de descendants appartenant à une classe (lisse - dominante) et à une autre classe (ridée - récessive). Il s'est avéré qu'environ un quart des graines étaient ridées et les trois quarts étaient lisses.
Mendel a réalisé les mêmes croisements d'hybrides de première génération pour toutes les autres caractéristiques : couleur des graines, couleur des fleurs, etc. Il a vu que le rapport 3:1 était maintenu.
Mendel s'est croisé dans un sens (père avec un trait dominant, mère avec un trait récessif) et dans l'autre (père avec un trait récessif, mère avec un trait dominant). Dans le même temps, les résultats qualitatifs et quantitatifs de la transmission des traits entre générations étaient les mêmes. De là, nous pouvons conclure que les inclinations féminines et paternelles du trait contribuent de manière égale à l'héritage du trait chez la progéniture.
Le fait que dans la première génération le trait d'un seul parent apparaisse, nous appelons la loi d'uniformité des hybrides de première génération ou la loi de dominance.
Le fait qu'à la deuxième génération les caractéristiques de l'un des parents (dominant) et de l'autre (récessif) réapparaissent a permis à Mendel de suggérer que ce n'est pas le trait en tant que tel qui est hérité, mais la tendance à son développement (ce que nous appelons maintenant le gène). Il a également suggéré que chaque organisme contient une paire de telles inclinations pour chaque trait. Une seule des deux inclinations passe du parent au descendant. Le dépôt de chaque type (dominant ou récessif) passe au descendant avec une probabilité égale. Lorsqu'un descendant combine deux inclinations différentes (dominante et récessive), une seule d'entre elles apparaît (dominante, désignée par une lettre majuscule A). L'inclinaison récessive (désignée par la petite lettre a) ne disparaît pas chez l'hybride, puisqu'elle se manifeste comme un trait dans la génération suivante.
Puisque dans la deuxième génération apparaissait exactement le même organisme que l'organisme parent, Mendel a décidé que le dépôt d'un trait « n'est pas obscurci » ; lorsqu'il est combiné avec un autre, il reste tout aussi pur. Par la suite, il a été découvert que seulement la moitié de ses inclinations sont transmises par un organisme donné - les cellules sexuelles, appelées gamètes, ne portent qu'une des deux caractéristiques alternatives.
Les humains ont environ 5 000 caractéristiques morphologiques et biochimiques qui sont héritées de manière assez claire selon Mendel. À en juger par la division au cours de la deuxième génération, les inclinations alternatives d'un trait se sont combinées indépendamment les unes des autres. Autrement dit, un trait dominant pourrait apparaître dans des combinaisons telles que Ahh, aA Et AA, et récessif uniquement en combinaison ahh.
Répétons que Mendel a suggéré que ce n'est pas le trait qui est hérité, mais les inclinations du trait (gènes) et que ces inclinations ne se mélangent pas, c'est pourquoi cette loi est appelée la loi de la pureté des gamètes. Grâce à l'étude du processus d'héritage, il a été possible de tirer des conclusions sur certaines caractéristiques du matériel hérité, à savoir que les inclinaisons sont stables au fil des générations, conservent leurs propriétés, que les inclinaisons sont discrètes, c'est-à-dire qu'il n'y a qu'un seul état de le trait est déterminé, qu'il y en a deux, ils sont combinés au hasard, etc. d.
À l'époque de Mendel, on ne savait rien de la méiose, même si structure nucléaire les cellules le savaient déjà. Le fait que le noyau contienne une substance appelée nucléine n’est devenu connu que quelques années après la découverte des lois de Mendel, et cette découverte n’avait aucun lien avec lui.
Toutes les conclusions du matériel ci-dessus peuvent être formulées comme suit :
1) Chaque caractéristique héréditaire est déterminée par un facteur héréditaire distinct, un dépôt ; dans la vision moderne, ces inclinations correspondent aux gènes ;
2) Les gènes sont conservés sous leur forme pure sur plusieurs générations, sans perdre leur individualité : c'est la preuve de l'essentiel de la génétique : le gène est relativement constant ;
3) Les deux sexes participent également à la transmission de leurs propriétés héréditaires à leur progéniture ;
4) Reduplication d'un nombre égal de gènes et leur réduction en cellules germinales mâles et femelles ; cette position était une prédiction génétique de l'existence de la méiose ;
5) Les inclinations héréditaires sont appariées, l'une est maternelle, l'autre est paternelle ; l'un d'eux peut être dominant, l'autre récessif ; Cette position correspond à la découverte du principe de l'allélisme : un gène est représenté par au moins deux allèles.
Les lois de l'héritage comprennent la loi du fractionnement des caractéristiques héréditaires chez la progéniture d'un hybride et la loi de la combinaison indépendante des caractéristiques héréditaires. Ces deux lois reflètent le processus de transmission des informations héréditaires dans les générations cellulaires lors de la reproduction sexuée. Leur découverte fut la première preuve concrète de l’existence de l’hérédité en tant que phénomène.
Les lois de l'hérédité ont un contenu différent, et elles se formulent ainsi :
- La première loi est la loi de la détermination héréditaire discrète (génétique) des traits ; il sous-tend la théorie des gènes.
- La deuxième loi est la loi de la constance relative de l'unité héréditaire - le gène.
- La troisième loi est la loi de l'état allélique du gène (dominance et récessivité).
Le fait que les lois de Mendel soient liées au comportement des chromosomes au cours de la méiose a été découvert au début du XXe siècle lors de la redécouverte des lois de Mendel par trois groupes de scientifiques indépendants les uns des autres. Comme vous le savez déjà, la particularité de la méiose est que le nombre de chromosomes dans une cellule est réduit de moitié ; les chromosomes peuvent changer de partie au cours de la méiose. Cette caractéristique caractérise la situation du cycle de vie chez tous les eucaryotes.
Afin de tester l'hypothèse de l'hérédité des inclinations sous cette forme, comme nous l'avons déjà dit, Mendel a également croisé les descendants de la première génération, qui avaient des graines jaunes avec du vert parental (récessif). Il a appelé le croisement un organisme récessif analysant. En conséquence, il a obtenu un partage un pour un : ( Ahh X ahh = Ahh + Ahh + ahh + ahh). Ainsi, Mendel a confirmé l'hypothèse selon laquelle dans le corps de la première génération se trouvent les caractéristiques des traits de chacun des parents dans un rapport de 1 pour 1. Mendel a appelé l'état où les deux caractéristiques des traits sont les mêmes homozygotes, et quand ils sont différents - hétérozygotes.

Mendel a pris en compte les résultats obtenus sur des milliers de graines, c'est-à-dire qu'il a mené des études statistiques reflétant un schéma biologique. Les lois mêmes qu’il a découvertes s’appliqueront également à d’autres eucaryotes, comme les champignons. On voit ici des champignons dans lesquels les quatre spores produites par une seule méiose restent dans une coquille commune. L'analyse du croisement chez de tels champignons conduit au fait que dans une coquille il y a 2 spores avec une caractéristique d'un parent et deux avec une caractéristique de l'autre. Ainsi, la division 1:1 dans un croisement d'analyse reflète le modèle biologique de division des inclinations d'un caractère à chaque méiose, qui ressemblera à un modèle statistique si toutes les spores sont mélangées.
Le fait que les parents avaient des états différents d'un trait suggère que les inclinations au développement de ce trait peuvent d'une manière ou d'une autre changer. Ces changements sont appelés mutations. Les mutations peuvent être neutres : forme des cheveux, couleur des yeux, etc. Certaines mutations entraînent des changements qui perturbent le fonctionnement normal de l’organisme. Il s'agit des pattes courtes chez les animaux (bovins, moutons, etc.), de l'absence de yeux et d'ailes chez les insectes, de l'absence de poils chez les mammifères, du gigantisme et du nanisme.
Certaines mutations peuvent être inoffensives, comme la pilosité chez l'homme, bien que tous les primates aient des cheveux. Mais parfois, il y a des changements dans l'intensité de la pilosité sur le corps et chez les personnes. N.I. Vavilov a appelé ce phénomène la loi des séries homologiques de variabilité héréditaire : c'est-à-dire qu'un trait typique d'une seule des deux espèces apparentées peut être trouvé avec une certaine fréquence chez les individus d'une espèce apparentée.
Cette diapositive montre que les mutations peuvent être assez visibles, on voit la famille noire dans laquelle il est né homme noir blanc- albinos. Ses enfants sont susceptibles d'être pigmentés, puisque cette mutation est récessive et que sa fréquence d'apparition est faible.

Nous avons déjà parlé de signes qui se manifestent pleinement. Mais cela n’est pas vrai pour tous les signes. Par exemple, le phénotype des hétérozygotes peut être intermédiaire entre les traits dominants et récessifs des parents. Ainsi, la couleur du fruit des aubergines de la première génération passe du bleu foncé à un violet moins intense. Dans le même temps, dans la deuxième génération, la répartition selon la présence de couleur est restée 3:1, mais si l'on prend en compte l'intensité de la couleur, la répartition est devenue 1:2:1 (couleur bleu foncé - AA, violet - 2 Ahh et blanc - ahh, respectivement) Dans ce cas, il est clair que la manifestation du trait dépend de la dose de l'allèle dominant. La division du phénotype correspond à la division du génotype : classes AA, Ahh Et ahh, dans un rapport de 1:2:1.

Soulignons une fois de plus le rôle de Mendel dans le développement de la science. Personne avant lui n’avait pensé que la création de signes pouvait exister. On croyait qu'à l'intérieur de chacun de nous il y avait un petit homme, en lui il y avait un autre petit homme, etc. La conception a quelque chose à voir avec son apparition, mais selon le mécanisme, le petit homme prêt est déjà présent dès le début de sa croissance. C'étaient les idées dominantes, qui, bien sûr, présentaient un inconvénient - selon cette théorie, avec un grand nombre de générations, l'homoncule aurait dû s'avérer plus petit qu'une particule élémentaire, mais ils ne savaient pas encore à propos des particules J.
Comment Mendel savait-il quel trait était dominant et lequel était récessif ? Il ne savait rien de tel, il a juste adopté un principe d'organisation de l'expérience. Heureusement, les signes qu’il observait étaient différents : hauteur, taille, couleur des fleurs, couleur des haricots, etc. Il ne disposait pas d’un modèle a priori du mécanisme d’hérédité ; il le dérivait de l’observation de la transmission d’un trait au fil des générations. Autre particularité de sa méthode. Il a constaté que la proportion d'individus présentant un trait récessif dans la deuxième génération représente un quart de l'ensemble de la progéniture. Autrement dit, la probabilité qu'un pois donné soit vert est de 1/4. Disons que nous avons en moyenne 4 pois dans une cosse. Chaque cosse (la progéniture de deux et seulement deux parents) contiendra-t-elle 1 pois vert et 3 pois jaunes ? Non. Par exemple, la probabilité qu’il y ait 2 pois verts est de 1/4 x 1/4 = 1/16, et que les quatre soient verts est de 1/256. Autrement dit, si vous prenez un tas de haricots contenant quatre pois chacun, alors un pois sur 256 aura des traits récessifs, c'est-à-dire verts. Mendel a analysé la progéniture de nombreux couples identiques de parents. On a parlé de croisement parce qu'ils montrent que les lois de Mendel apparaissent comme des lois statistiques, mais sont basées sur un modèle biologique - 1:1. C'est-à-dire les gamètes différents types dans CHAQUE méiose, les hétérozygotes se forment dans un rapport égal - 1:1, et les modèles se manifestent statistiquement, puisque les descendants de centaines de méioses sont analysés - Mendel a analysé plus de 1000 descendants dans chaque type de croisement.
Mendel a d'abord étudié l'héritage d'une paire de traits. Il se demande alors ce qui se passerait si deux paires de signes étaient observées simultanément. La figure ci-dessus, à droite, illustre une telle étude sur les paires de signes de pensée – la couleur des pois et la forme des pois.
Les parents d'un type ont produit des pois jaunes et ronds lors de l'autopollinisation. Les parents d'un autre type ont produit des pois verts et ridés lors de l'autopollinisation. Dans la première génération, tous les pois étaient jaunes et de forme ronde. La division qui en résulte dans la deuxième génération peut être facilement examinée à l’aide du réseau Penet. Nous avons obtenu un fractionnement selon les caractéristiques 9:3:3:1 (jaune et rond : jaune et ridé : vert et rond : vert et ridé). Le fractionnement pour chaque paire de caractéristiques se produit indépendamment les uns des autres. Le rapport 9zhk + 3zhm + 3zk + 1zm correspond à une combinaison indépendante des résultats de deux croisements (3zh + 1z) x (3k + 1m). Autrement dit, les caractéristiques de ces paires (couleur et forme) sont combinées indépendamment.
Comptons combien de classes phénotypiques différentes nous avons. Nous avions 2 classes phénotypiques : jaune et verte ; et selon une autre caractéristique, 2 classes phénotypiques : rondes et ridées. Et au total, il y aura 2*2=4 classes phénotypiques, ce que nous avons obtenu ci-dessus. Si l’on considère trois caractéristiques, alors il y aura 2 3 =8 classes phénotypiques. Mendel est allé jusqu'aux croisements dihybrides. Heureusement pour Mendel, la composition de tous les caractères était localisée sur différents chromosomes chez les pois, et il y a 7 paires de chromosomes chez les pois. Par conséquent, il s’est avéré qu’il a adopté des traits qui se sont combinés indépendamment chez la progéniture.
Les humains possèdent 23 paires de chromosomes. Si l'on considère un trait hétérozygote pour chaque chromosome, une personne peut avoir 2 23 ~ 8 * 10 6 classes phénotypiques chez la progéniture d'un couple marié. Comme mentionné dans la première leçon, chacun de nous contient environ 1 différence entre les chromosomes de notre père et de notre mère pour 1000 positions, soit un total d'environ un million de différences entre les chromosomes de notre père et de notre mère. Autrement dit, chacun de nous est le descendant d'un croisement d'un million d'hybrides, dans lequel le nombre de classes phénotypiques est de 2 1000 000. En pratique, ce nombre de classes phénotypiques n'est pas réalisé dans la progéniture d'un couple, car nous n'avons que 23 chromosomes, et non un million. Il s'avère que 8 * 10 6 est la limite inférieure de la diversité possible de la progéniture d'un couple marié donné. Sur cette base, vous pouvez comprendre qu'il ne peut pas y avoir deux personnes absolument identiques. La probabilité de mutation d'un nucléotide donné dans l'ADN au cours d'une génération est d'environ 10 -7 - 10 -8, c'est-à-dire que pour l'ensemble du génome (3 * 10 9), il y aura environ 100 changements de novo entre le parent et l'enfant. Et les différences totales entre la moitié paternelle de votre génome et la moitié maternelle sont d'environ 1 000 000. Cela signifie que les anciennes mutations de votre génome sont beaucoup plus fréquentes que les nouvelles mutations (10 000 fois).
Mendel a également réalisé un croisement analytique - croisement avec un homozygote récessif. Chez un descendant de la première génération, la combinaison de gènes a la forme AAB b. Si vous le croisez avec un représentant avec un ensemble de gènes complètement récessifs ( aabb), nous obtiendrons alors quatre classes possibles, qui seront dans le rapport 1:1:1:1, contrairement au croisement évoqué ci-dessus, où nous avons obtenu une répartition de 9:3:3:1.

Vous trouverez ci-dessous quelques critères statistiques : quels ratios de nombres doivent être considérés comme cohérents avec ce qui est attendu, par exemple 3 : 1. Par exemple, pour 3: 1 - sur quatre cents pois, il est peu probable qu'il en résulte exactement 300 à 100. S'il s'avère, par exemple, 301 à 99, alors ce rapport peut probablement être considéré comme égal à 3 pour 1. Mais 350 pour 50 n’est probablement pas égal à 3 pour 1.
La statistique du test du chi carré (χ 2) est utilisée pour tester l'hypothèse selon laquelle la distribution observée correspond à celle attendue. Cette lettre grecque se prononce en russe par « chi » et en anglais par « chi » (chi).
La valeur de χ 2 est calculée comme la somme des carrés des écarts des valeurs observées par rapport à la valeur attendue, divisée par la valeur attendue. Ensuite, à l'aide d'un tableau spécial pour une valeur donnée de χ 2, trouvez la probabilité qu'une telle différence entre la valeur observée et attendue soit aléatoire. Si la probabilité est inférieure à 5 %, alors l'écart est considéré comme non aléatoire (le chiffre de cinq pour cent a été choisi d'un commun accord).

Un trait héréditaire prédéterminé se manifestera-t-il toujours ? Après tout, cette hypothèse par défaut sous-tend l’interprétation des données obtenues par Mendel.
Il s’avère que cela peut dépendre de plusieurs raisons. Il existe un tel trait héréditaire chez l'homme : les six doigts. Bien que, comme tous les vertébrés, nous ayons normalement cinq doigts.
La probabilité de manifestation de l'inclination d'un trait sous la forme d'un trait observable (ici - six doigts) peut être inférieure à 100 %. Sur la photo, l'homme a 6 orteils aux deux pieds. Et son jumeau ne présentera pas forcément ce signe. La proportion d'individus possédant un génotype donné et présentant le phénotype correspondant était appelée pénétrance (ce terme a été introduit par le généticien russe Timofeev-Resovsky).
Dans certains cas, le sixième chiffre peut simplement être marqué par une excroissance cutanée. Timofeev-Resovsky a proposé d'appeler le degré d'expression d'un trait une expressivité individuelle.
Le lien qui n'est pas à 100 % entre le génotype et le phénotype est particulièrement clair lors de l'étude de vrais jumeaux. Leur constitution génétique est la même et leurs caractéristiques coïncident à des degrés divers. Vous trouverez ci-dessous un tableau montrant la coïncidence des caractéristiques des jumeaux identiques et non identiques. Diverses maladies sont considérées comme des signes dans ce tableau.
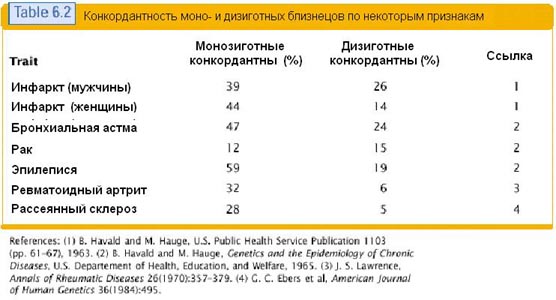
Un trait présent chez la plupart des individus conditions naturelles l'habitat est appelé type sauvage. Le trait le plus courant est souvent dominant. Une telle connexion peut avoir une signification adaptative bénéfique pour l’espèce. Chez l’homme, les caractéristiques dominantes sont par exemple les cheveux noirs, les yeux foncés, les cheveux bouclés. À propos, puisque les gènes correspondants se trouvent sur des chromosomes différents, vous pouvez obtenir un homme noir aux cheveux bouclés qui sera blond - rien ne l'interdit.
Pourquoi se fait-il que dans un croisement monohybride, trois classes génotypiques dans la descendance de deuxième génération correspondent dans certains cas à trois classes phénotypiques (aubergines bleues violettes et blanches), et dans d'autres cas à deux classes (pois jaunes ou verts) ? Pourquoi la manifestation d’un trait dominant est-elle incomplète dans un cas et complète dans l’autre ? Une analogie peut être établie avec le film photographique. Selon la quantité de lumière, le cadre peut s'avérer complètement transparent, gris ou complètement noir. C'est la même chose avec les gènes. Par exemple, le maïs possède un gène Y, qui détermine la formation de vitamine A. Lorsque la dose du gène Y par cellule passe de un à trois, l'activité de l'enzyme qu'il code change de manière linéaire et, dans ce cas, la formation de vitamine A et la couleur du grain augmente. (Dans le maïs, la partie principale du grain est l'endosperme. Chaque cellule de l'endosperme contient trois génomes - deux de la mère et un du père). Autrement dit, de nombreux caractères dépendent quantitativement de la dose d’allèle. Plus il y a de copies d’un allèle du type souhaité, plus la valeur du trait qu’il contrôle est grande. Cette connexion est constamment utilisée en biotechnologie.

Mendel aurait pu ne pas découvrir ses lois. Les recherches sur le pois ont permis à Mendel de découvrir ses lois, car le pois est une plante autogame, et donc homozygote sans contrainte. Lors de l'autopollinisation, la proportion d'hétérozygotes diminue proportionnellement à deux puissances du nombre de générations. Ce fut la chance de Mendel : si la proportion d’hétérozygotes avait été importante, aucune tendance n’aurait été observée. Lorsqu'il a ensuite pris des pollinisateurs croisés, les schémas se sont rompus, ce qui a grandement contrarié Mendel car il pensait avoir découvert quelque chose de privé. Il s’est avéré que non.

Nous avons parlé ci-dessus de l’héritage de traits qualitatifs, mais la plupart des traits sont généralement quantitatifs. Leur contrôle génétique est assez complexe. Les caractéristiques quantitatives sont décrites par la valeur moyenne de la caractéristique et la plage de variation, appelée norme de réaction. La valeur moyenne et la norme de réaction sont des indicateurs spécifiques à l'espèce qui dépendent à la fois du génotype et des conditions environnementales. Par exemple, l'espérance de vie humaine. Même si la Bible dit que les prophètes ont vécu 800 ans, il est désormais clair que personne ne vit plus de 120 à 150 ans. Et une souris, par exemple, vit deux ans, même si elle est aussi un mammifère. Notre taille, notre poids, ce sont tous des signes quantitatifs. Il n'y a personne de 3 à 4 mètres de haut, bien qu'il y ait des éléphants, par exemple. Chaque espèce a sa propre moyenne pour chaque caractère quantitatif et sa propre plage de variation.

Des modèles d'hérédité sont découverts dans l'étude des traits qualitatifs.
La plupart de nos traits sont quantitatifs.
Les valeurs des traits dans un échantillon représentatif d'individus d'une espèce donnée sont caractérisées par une certaine moyenne et une certaine ampleur de variation, appelée norme de réaction et dépend à la fois du génotype et des conditions de formation du trait.
Sujet 4.2 Modèles de base
hérédité
Terminologie 1.Alternative– des signes contrastés. 2. Des lignes épurées– des plantes en rangée dont aucun dédoublement n'est observé lors de l'autopollinisation. 3. Méthode hybridologique– l'obtention d'une progéniture hybride et son analyse. 4. Parents– R.5. Mâles – ♂. 6. Femelles – ♀. 7. Croisement– X.8. Hybrides F 1, F 2, Fn. 9. Monohybride– croisement d’individus présentant un trait contrasté. Modèles d'héritage des traits Les modèles quantitatifs d'héritage des traits ont été découverts par le botaniste amateur tchèque G. Mendel. S'étant fixé pour objectif de découvrir les modèles d'hérédité des traits, il s'est tout d'abord concentré sur le choix de l'objet d'étude. Pour ses expériences, G. Mendel a choisi les pois, des variétés qui différaient clairement les unes des autres par un certain nombre de caractéristiques. L'un des points les plus importants de tous les travaux a été de déterminer le nombre de caractéristiques permettant de distinguer les plantes croisées. G. Mendel s'est d'abord rendu compte qu'en commençant par le cas le plus simple - les différences entre les parents sur un seul trait et en compliquant progressivement la tâche, on peut espérer démêler tout l'enchevêtrement des modèles de transmission des traits de génération en génération, c'est-à-dire leur héritage. Ici, la stricte nature mathématique de sa pensée se révèle. C'est cette approche qui a permis à G. Mendel de planifier clairement la complication ultérieure des expériences. À cet égard, Mendel était au-dessus de tous les biologistes de son temps. Un autre caractéristique importante ses recherches consistaient à choisir pour ses expériences des organismes appartenant à des lignées pures, c'est-à-dire de telles plantes, dans un certain nombre de générations dont, lors de l'autopollinisation, aucune ségrégation selon le trait étudié n'a été observée. Non moins important est qu'il a observé l'héritage des alternatives, c'est-à-dire des traits contrastés. Par exemple, les fleurs d'une plante étaient violettes et les fleurs d'une autre étaient blanches, la croissance de la plante était haute ou courte, les haricots étaient lisses ou ridés, etc. En comparant les résultats d'expériences et de calculs théoriques, G. Mendel a particulièrement souligné le caractère statistique moyen des modèles qu'il a découverts. Ainsi, la méthode de croisement d'individus qui diffèrent par des caractéristiques alternatives, c'est-à-dire l'hybridation, suivie d'une prise en compte stricte de la répartition des caractéristiques parentales dans la progéniture, est dite hybridiologique. Les schémas de transmission des traits, identifiés par G. Mendel et confirmés par de nombreux biologistes sur des objets variés, sont actuellement formulés sous la forme de lois qui régissent tous caractère général. Loi d'uniformité de la première génération d'hybrides Traversée monohybride. Pour illustrer la loi d'uniformité de la première génération - la première loi de Mendel, reproduisons ses expériences de croisement monohybride de plants de pois. Le monohybride est le croisement de deux organismes qui diffèrent l'un de l'autre par une paire de caractéristiques alternatives. Par conséquent, avec un tel croisement, il est possible de retracer des modèles de transmission de seulement deux variantes du trait, dont le développement est déterminé par une paire de gènes alléliques. Par exemple, un signe est la couleur des graines, les options sont jaune ou verte. Toutes les autres caractéristiques caractéristiques de ces organismes ne sont pas prises en compte. Si vous croisez des plants de pois avec des graines jaunes et vertes, tous les descendants hybrides résultants auront des graines jaunes. La même image est observée lors du croisement de plantes à graines lisses et ridées - toutes les graines des hybrides seront lisses. Par conséquent, dans un hybride de première génération, un seul de chaque paire de caractères alternatifs apparaît. Le deuxième signe semble disparaître et n'apparaît pas. Mendel a appelé la prédominance du trait de l'un des parents une dominance hybride. Un trait qui apparaît dans un hybride de première génération et supprime le développement d'un autre trait est appelé dominant, opposé, c'est-à-dire le trait supprimé est récessif. Un trait dominant est généralement désigné par une lettre majuscule (A) et un trait récessif par une lettre minuscule (a). Mendel a utilisé dans ses expériences des plantes appartenant à différentes lignées pures, ou variétés, dont les descendants étaient semblables à leurs parents sur une longue série de générations. Par conséquent, dans ces plantes, les deux gènes alléliques sont identiques. Ainsi, si le génotype d'un organisme contient deux gènes alléliques identiques, c'est-à-dire deux séquences nucléotidiques génétiques absolument identiques, un tel organisme est appelé homozygote. Un organisme peut être homozygote pour les gènes dominants (AA) ou récessifs (aa). Si les gènes alléliques diffèrent les uns des autres par la séquence nucléotidique, par exemple l'un est dominant et l'autre récessif (Aa), un tel organisme est appelé hétérozygote. La première loi de Mendel est aussi appelée loi de dominance ou d'uniformité, puisque tous les individus de la première génération ont la même manifestation d'un trait inhérent à l'un des parents. Il est formulé ainsi : Lors du croisement de deux organismes appartenant à des lignées pures différentes (deux homozygotes), différant l'un de l'autre par une paire de traits alternatifs, toute la première génération d'hybrides (F 1) sera uniforme et portera le trait d'un parent. En termes de couleur, Mendel a établi que le rouge ou le noir domineraient le blanc, le rose et le gris d'intensités variables étant des couleurs intermédiaires. Mendel a proposé des désignations graphiques pour les personnages : P – parents, ♂ – homme, ♀ – femme,, - gamètes, X - croisement, F 1, F 2, F n - progéniture. La première loi de Mendel est présentée dans la figure 1.
![]()
Figure 1. Première loi de Mendel
Tous les descendants ont la même couleur intermédiaire, ce qui ne contredit pas la première loi de Mendel.
Questions de contrôle
1. Matériel biologique de Mendel. 2. Signes alternatifs dans les expériences de Mendel. 3. Des lignes épurées et leur définition. 4. L'essence de la méthode hybridiologique. 5. Croisement monohybride. 6. Traits dominants et récessifs. 7. Gènes alléliques. 8. Première loi de Mendel. Loi d'uniformité.
Thème 4.2.1 Dominance incomplète des gènes
Terminologie 1. Gènes alléliques– des gènes situés dans des locus identiques de chromosomes homologues. 2. Trait dominant– supprimer le développement d’un autre. 3. Trait récessif- supprimé. 4. Homozygote- un zygote qui possède les mêmes gènes. 5. Hétérozygote- un zygote avec des gènes différents. 6. Diviser– divergence des traits chez la progéniture. 7. Traverser- chevauchement des chromosomes. A l'état hétérozygote, le gène dominant ne supprime pas toujours complètement la manifestation du gène récessif. Dans certains cas, l'hybride F 1 ne reproduit pas complètement l'un des caractères parentaux et l'expression du trait est de nature intermédiaire avec un biais plus ou moins grand vers un état dominant ou récessif. Mais tous les individus de cette génération présentent une uniformité dans ce trait. Le caractère intermédiaire de l'héritage dans le schéma précédent ne contredit pas la première loi de Mendel, puisque tous les descendants de F 1 sont uniformes. Dominance incomplète- un phénomène répandu. Elle a été découverte en étudiant l'héritage de la couleur des fleurs chez les mufliers, la structure des plumes des oiseaux, la couleur de la laine des bovins et des moutons, les caractéristiques biochimiques chez l'homme, etc. Allélisme multiple. Jusqu'à présent, nous avons analysé des exemples dans lesquels le même gène était représenté par deux allèles – dominant (A) et récessif (a). Ces deux conditions génétiques sont dues à une mutation. Un gène peut muter à plusieurs reprises. En conséquence, plusieurs variantes de gènes alléliques apparaissent. L’ensemble de ces gènes alléliques, qui déterminent la diversité des variantes du trait, est appelé série de gènes alléliques. L'apparition d'une telle série en raison de mutations répétées d'un gène est appelée allélisme multiple ou allélomorphisme multiple. Le gène A peut muter dans les états a 1, a 2, a 3 et n. Le gène B, situé dans un autre locus, est dans l'état b 1, b 2, b 3, b n. Par exemple, chez la mouche drosophile, on connaît une série d'allèles pour le gène de la couleur des yeux, composé de 12 membres : rouge, corail, cerise, abricot, etc. au blanc, déterminé par un gène récessif. Les lapins ont une série d’allèles multiples pour la couleur du pelage. Cela provoque le développement d’une couleur unie ou un manque de pigmentation (albinisme). Les membres d’une même série d’allèles peuvent entretenir des relations dominantes-récessives différentes les uns avec les autres. Il convient de rappeler que le génotype des organismes diploïdes ne peut contenir que deux gènes issus d'une série d'allèles. Les allèles restants de ce gène dans différentes combinaisons sont inclus par paires dans les génotypes d'autres individus de cette espèce. Ainsi, l'allélisme multiple caractérise la diversité du pool génétique, c'est-à-dire la totalité de tous les gènes qui composent les génotypes d'un certain groupe d'individus ou d'une espèce entière. En d’autres termes, l’allélisme multiple est un trait d’espèce et non un trait individuel. Deuxième loi de Mendel - Loi de ségrégation Si les descendants de la première génération, identiques dans le trait étudié, sont croisés entre eux, alors à la deuxième génération les traits des deux parents apparaissent dans un certain rapport numérique : 3/4 des individus auront un trait dominant, 1 /4 en aura un récessif. Selon le génotype en F 2, il y aura 25 % d'individus homozygotes pour les allèles dominants, 50 % des organismes seront hétérozygotes et 25 % de la progéniture sera des organismes homozygotes pour les allèles récessifs. Le phénomène dans lequel le croisement d'individus hétérozygotes conduit à la formation de descendants, dont certains portent un trait dominant et d'autres un trait récessif, est appelé ségrégation. Par conséquent, la ségrégation est la répartition des traits dominants et récessifs parmi la progéniture dans un certain rapport numérique. Le caractère récessif ne disparaît pas chez les hybrides de première génération, mais est seulement supprimé et apparaît dans la deuxième génération hybride. Ainsi, la deuxième loi de Mendel (voir Fig. 2) peut être formulée comme suit : lorsque deux descendants de la première génération sont croisés (deux hétérozygotes), dans la deuxième génération on observe une scission dans un certain rapport numérique : par phénotype 3:1, par génotype 1 : 2:1.
Figure 2. Deuxième loi de Mendel
Avec une dominance incomplète dans la progéniture des hybrides F 2, la ségrégation du génotype et du phénotype coïncide (1 : 2 : 1). Loi de la pureté des gamètes Cette loi reflète l'essence du processus de formation des gamètes lors de la méiose. Mendel a suggéré que les facteurs héréditaires (gènes) ne se mélangent pas lors de la formation des hybrides, mais restent inchangés. Dans le corps de l'hybride F, issu du croisement de parents qui diffèrent par des caractéristiques alternatives, les deux facteurs sont présents - dominant et récessif. Le facteur héréditaire dominant se manifeste sous la forme d'un trait, tandis que le facteur récessif est supprimé. Le lien entre les générations lors de la reproduction sexuée s'effectue à travers les cellules germinales - les gamètes. Par conséquent, il faut supposer que chaque gamète ne porte qu’un seul facteur d’une paire. Puis, lors de la fécondation, la fusion de deux gamètes, chacun porteur d'un facteur héréditaire récessif, va conduire à la formation d'un organisme au trait récessif qui se manifeste de manière phénotypique. La fusion de gamètes porteurs d'un facteur dominant, ou de deux gamètes dont l'un contient un facteur dominant et l'autre un facteur récessif, conduira au développement d'un organisme doté d'un trait dominant. Ainsi, l'apparition dans la deuxième génération (F 2) d'un trait récessif de l'un des parents (P) ne peut se produire que si deux conditions sont remplies : 1. Si chez les hybrides les facteurs héréditaires restent inchangés. 2. Si les cellules germinales ne contiennent qu'un seul facteur héréditaire issu d'une paire allélique. Mendel a expliqué la division des caractéristiques de la progéniture lors du croisement d'individus hétérozygotes par le fait que les gamètes sont génétiquement purs, c'est-à-dire ne portent qu'un seul gène d'une paire allélique. La loi de la pureté des gamètes peut être formulée comme suit : lors de la formation des cellules germinales, un seul gène d'une paire allélique (de chaque paire allélique) pénètre dans chaque gamète. La preuve cytologique de la loi de pureté des gamètes est le comportement du chromosome en méiose : dans la première division méiotique, les chromosomes homologues se retrouvent dans des cellules différentes, et dans l'anaphase de la seconde, les chromosomes filles, qui, en raison du croisement, peuvent contiennent différents allèles du même gène. On sait que chaque cellule du corps possède exactement le même ensemble diploïde de chromosomes. Deux chromosomes homologues contiennent deux gènes alléliques identiques. La formation de gamètes génétiquement « purs » est illustrée dans le diagramme de la figure 3.

Figure 3. Formation de gamètes « purs »
Lorsque les gamètes mâles et femelles fusionnent, un hybride se forme et possède un ensemble diploïde de chromosomes (voir Fig. 4).

Figure 4. Formation hybride
Comme le montre le diagramme, le zygote reçoit la moitié des chromosomes du corps du père et l’autre moitié de celui de la mère. Lors de la formation des gamètes chez un hybride, les chromosomes homologues se retrouvent également dans différentes cellules lors de la première division méiotique (voir Fig. 5).
Figure 5. Formation de deux types de gamètes
Deux variétés de gamètes se forment selon une paire allélique donnée. Ainsi, la base cytologique de la loi de pureté des gamètes, ainsi que la division des caractéristiques de la progéniture lors du croisement monohybride, est la divergence des chromosomes homologues et la formation de cellules haploïdes lors de la méiose. Croix d'analyse La méthode hybridologique d'étude de l'hérédité développée par Mendel permet d'établir si un organisme possédant un phénotype dominant pour le gène étudié est homozygote ou hétérozygote. La race est-elle pure ? Pour ce faire, un individu de génotype inconnu est croisé avec un organisme homozygote pour l'allèle récessif et possédant un phénotype récessif. Si l'individu dominant est homozygote, la progéniture issue d'un tel croisement sera uniforme et aucune division ne se produira (voir Fig. 6).

Figure 6. Croisement d'individus dominants.
Le tableau sera différent si l'organisme étudié est hétérozygote (voir Fig. 7).

Figure 7. Croisement d'individus hétérozygotes.
Le clivage se produira dans un rapport 1:1 selon le phénotype. Ce résultat de croisement est la preuve de la formation de deux variétés de gamètes chez l'un des parents, à savoir son hétérozygotie n'est pas une race pure (voir Fig. 8).

Figure 8. La ségrégation se produira dans un rapport de 1 : 1 selon le phénotype.
Questions de contrôle
1. Domination incomplète et sa manifestation dans la nature. 2. L'essence de l'allélisme multiple. 3. II-Loi de Mendel. La loi du fractionnement. 4. La loi de la pureté des gamètes. 5. Preuve cytologique de la loi de pureté des gamètes. 6. Analyser le croisement, son essence et sa signification.
Thème 4.2.2 III Loi de Mendel - la loi de l'indépendance
combinant des fonctionnalités
Terminologie 1. Croisement dihybride– croisement pour deux caractéristiques contrastées. 2. Organismes dihétérozygotes– les organismes hétérozygotes pour deux paires de gènes alléliques. 3. Grille à pannet– méthode graphique de calcul des résultats de croisement. 4. Recombinaison– recombinaison de fonctionnalités. 5. Traverser– l’apparition de nouveaux caractères lors du chevauchement des chromosomes. 6. Morganide– la distance entre les gènes. Croisement dihybride et polyhybride Les organismes diffèrent les uns des autres à bien des égards. Les modèles d'hérédité de deux ou plusieurs paires de caractères alternatifs peuvent être établis par croisement dihybride ou polyhybride. Pour le croisement dihybride, Mendel a utilisé des plants de pois homozygotes qui différaient par deux paires de caractéristiques : la couleur des graines (jaune et verte) et la forme des graines (lisse et ridée). Les dominantes étaient la couleur jaune (A) et la forme lisse des graines (B). Chaque plante produit une variété de gamètes selon les allèles étudiés. Lorsque les gamètes fusionnent, tous les descendants seront uniformes (voir Fig. 9).
Figure 9. Fusion de gamètes
Les organismes hétérozygotes pour deux paires de gènes alléliques sont appelés dihétérozygotes. Lorsque les gamètes se forment dans un hybride, à partir de chaque paire de gènes alléliques, un seul entre dans le gamète, et en raison du caractère aléatoire de la divergence des chromosomes paternels et maternels lors de la première division de la méiose, le gène A peut se retrouver dans le le même gamète avec le gène B ou le gène b, tout comme le gène a peut se combiner en un seul gamète avec le gène B ou avec le gène b (voir Fig. 10).

Figure 10. Formation de gamètes chez un hybride
Tableau 1.
Traitement des résultats des croisements dihybrides
| UN B | AABB | AABb | AaBB | AaBb |
| Un B | AABb | AAbb | AaBb | Aabb |
| un B | AaBB | AaBb | aaBB | aaBb |
| un B | AaBb | Aabb | aaBb | aabb |
Les lois d'hérédité des caractères de G. Mendel décrivent les principes fondamentaux de la transmission des caractères héréditaires des organismes parents à leurs enfants ; ces principes sous-tendent la génétique classique. Ces lois ont été découvertes par Mendel à la suite du croisement d'organismes (dans ce cas, des plantes) avec différents génotypes. Habituellement, une règle et deux lois sont décrites.
Règle d'uniformité hybride de première génération
En croisant des pois de semence avec des traits stables - fleurs violettes et blanches, Mendel a remarqué que les hybrides qui ont émergé étaient tous avec des fleurs violettes, il n'y en avait pas une seule blanche parmi eux. Mendel a répété ses expériences plus d'une fois et a utilisé d'autres signes. Par exemple, s'il croisait des pois avec des graines jaunes et vertes, la progéniture avait des graines jaunes ; lorsqu'il croisait des pois avec des graines lisses et ridées, la progéniture avait des graines lisses. La progéniture des plantes hautes et courtes était grande.
Donc, les hybrides de première génération acquièrent toujours l'une des caractéristiques parentales. Un trait (plus fort, dominant) en supprime toujours un autre (plus faible, récessif). Ce phénomène est appelé domination totale.
Si nous appliquons la règle ci-dessus à une personne, par exemple en utilisant l'exemple des yeux bruns et bleus, alors elle s'explique comme suit. Si chez un parent homozygote, les deux gènes du génome déterminent la couleur des yeux marron (nous désignons un génotype tel que AA), et chez un autre, également homozygote, les deux gènes déterminent la couleur des yeux bleus (nous désignons ce génotype par ahh), alors les gamètes haploïdes qu'ils produisent porteront toujours soit le gène UN, ou UN(voir schéma ci-dessous).
Schéma de transfert de caractère lors du croisement d'organismes homozygotes
Alors tous les enfants auront le génotype Ahh, mais tout le monde aura les yeux bruns car le gène des yeux bruns est dominant sur le gène des yeux bleus.
Voyons maintenant ce qui se passe si des organismes hétérozygotes (ou hybrides de première génération) sont croisés. Dans ce cas, cela arrivera fractionnement des caractères dans certaines relations quantitatives.
La loi de ségrégation des caractères, ou première loi de Mendel
Si hétérozygote les descendants de la première génération, identiques dans le trait étudié, sont croisés entre eux, puis dans la deuxième génération les traits des deux parents apparaissent dans un certain rapport numérique : 3/4 des individus auront un trait dominant, 1/4 auront un trait récessif(voir schéma ci-dessous).

Modèle d'héritage des traits lors du croisement d'organismes hétérozygotes
Le phénomène dans lequel le croisement d'individus hétérozygotes conduit à la formation de descendants, dont certains portent un trait dominant et d'autres un trait récessif, est appelé scission. Comme nous le comprenons, le trait récessif n’a pas disparu chez les hybrides de première génération, mais a seulement été supprimé et est apparu dans la deuxième génération hybride. Mendel a été le premier à comprendre que lors de la formation des hybrides, les facteurs héréditaires ne sont pas mélangés ou « érodés », mais restent inchangés. Dans un organisme hybride, les deux facteurs (gènes) sont présents, mais seul le facteur héréditaire dominant se manifeste sous forme de trait.
Le lien entre les générations lors de la reproduction sexuée s'effectue à travers les cellules germinales, chaque gamète ne porte qu'un seul facteur du couple. La fusion de deux gamètes, dont chacun porte un facteur héréditaire récessif, conduira à l'apparition d'un organisme à caractère récessif. La fusion de gamètes, dont chacun porte un facteur dominant, ou de deux gamètes dont l'un contient un facteur dominant et l'autre un facteur récessif, conduit au développement d'un organisme avec un trait dominant.
Mendel a expliqué la division lors du croisement d'individus hétérozygotes par le fait que les gamètes ne portent qu'un seul gène d'une paire allélique ( loi de la pureté des gamètes). En effet, cela n'est possible que si les gènes restent inchangés et que les gamètes ne contiennent qu'un seul gène d'une paire. Il est pratique d'étudier les relations entre les entités en utilisant ce que l'on appelle le réseau de Punnett :
| Un (0,5) | une (0,5) | |
| Un (0,5) | AA (0,25) | Aa (0,25) |
| une (0,5) | Aa (0,25) | aa (0,25) |
En raison de la probabilité statistique, avec un nombre suffisamment grand de gamètes dans la progéniture, 25 % des génotypes seront homozygotes dominants, 50 % seront hétérozygotes, 25 % seront homozygotes récessifs, c'est-à-dire qu'une relation mathématique s'établit : 1 AA:2Ahh:1ahh. En conséquence, selon le phénotype, la progéniture de la deuxième génération lors d'un croisement monohybride est répartie dans un rapport de 3:1 - 3 parties d'individus avec un trait dominant, 1 partie d'individus avec un trait récessif.
Il ne faut pas oublier que la répartition des gènes et leur entrée dans les gamètes sont probabilistes. L'approche de Mendel pour l'analyse des descendants était quantitative et statistique : tous les descendants avec un état de trait donné (par exemple, les pois lisses ou ridés) étaient regroupés en un seul groupe, leur nombre était calculé, qui était comparé au nombre de descendants avec un autre état du trait (pois ridés). Cette analyse par paires a assuré le succès de ses observations. Dans le cas d'une personne, il peut être très difficile d'observer une telle répartition - il est nécessaire qu'un couple de parents ait au moins une douzaine d'enfants, ce qui est assez rare dans la société moderne. Il se peut donc que des parents aux yeux bruns donnent naissance à un seul enfant, et que cet enfant ait les yeux bleus, ce qui, à première vue, viole toutes les lois de la génétique. En même temps, si vous expérimentez avec la drosophile ou des souris de laboratoire, les lois mendéliennes sont assez faciles à observer.
Il faut dire que dans un certain sens, Mendel a eu de la chance: dès le début, il a choisi comme objet une plante appropriée: des pois colorés. S’il tombait par exemple sur des plantes comme la belle de nuit ou le muflier, le résultat serait imprévisible. Le fait est que chez les mufliers, les plantes hétérozygotes obtenues en croisant des plantes homozygotes à fleurs rouges et blanches ont des fleurs roses. De plus, aucun des allèles ne peut être qualifié de dominant ou de récessif. Ce phénomène peut s'expliquer par le fait que des processus biochimiques, en raison des performances différentes des allèles, ne conduisent pas nécessairement à des résultats alternatifs mutuellement exclusifs. Le résultat peut être intermédiaire, selon les caractéristiques du métabolisme dans organisme donné, dans lequel il existe toujours de nombreuses options, mécanismes de dérivation ou processus existants parallèles avec différentes manifestations externes.
Ce phénomène est appelé dominance incomplète ou codominance, cela se produit assez souvent, y compris chez l'homme. Un exemple est le système de groupes sanguins humains MN (notez au passage qu'il ne s'agit que d'un des systèmes ; il existe de nombreuses classifications de groupes sanguins). Landsteiner et Levin expliquaient autrefois ce phénomène par le fait que les globules rouges pouvaient porter à leur surface soit un antigène (M), soit un autre (N), soit les deux ensemble (MN). Si dans les deux premiers cas nous avons affaire à des homozygotes (MM et NN), alors à l'état hétérozygote (MN) les deux allèles se manifestent et les deux apparaissent (dominent), d'où le nom de codominance.
La loi de l'héritage indépendant des caractères, ou deuxième loi de Mendel
Cette loi décrit la répartition des caractéristiques sous ce qu'on appelle dihybride Et polyhybride croisement, c'est-à-dire lorsque les individus croisés diffèrent par deux ou plusieurs caractéristiques. Dans les expériences de Mendel, des plantes croisées différaient par plusieurs paires de traits, tels que : 1) des fleurs blanches et violettes et 2) des graines jaunes ou vertes. De plus, l'héritage de chaque trait suivait les deux premières lois, et les traits étaient combinés indépendamment les uns des autres. Comme prévu, la première génération après croisement avait un phénotype dominant à tous égards. La deuxième génération a suivi la formule 9:3:3:1, c'est-à-dire que 9/16 exemplaires étaient à fleurs violettes et pois jaunes, 3/16 étaient à fleurs blanches et pois jaunes, 3/16 autres étaient à fleurs violettes et vertes. pois et, enfin, 1/16 - avec des fleurs blanches et des pois verts. Cela s'est produit parce que Mendel a réussi à sélectionner des caractères dont les gènes étaient situés sur différents chromosomes de pois. La deuxième loi de Mendel n'est vraie que dans les cas où les paires de gènes analysées sont situées sur des chromosomes différents. Selon la règle de la fréquence des gamètes, les caractères sont combinés indépendamment les uns des autres, et s'ils se trouvent sur des chromosomes différents, l'héritage des caractères se produit indépendamment.
Les première et deuxième lois de Mendel sont universelles, mais il existe toujours des exceptions à la troisième loi. La raison en devient claire si l'on se souvient qu'il existe de nombreux gènes sur un chromosome (chez l'homme, de plusieurs centaines à un millier ou plus). Si les gènes sont sur le même chromosome, il peut y avoir héritage enchaîné. Dans ce cas, les caractéristiques sont transmises par paires ou par groupes. Les gènes situés sur le même chromosome sont appelés en génétique groupes d'embrayage. Le plus souvent, les traits déterminés par des gènes proches les uns des autres sur le chromosome sont transmis ensemble. De tels gènes sont appelés étroitement imbriqués. Dans le même temps, il arrive parfois que des gènes éloignés les uns des autres soient hérités de manière liée. La raison de ce comportement différent des gènes est un phénomène particulier échange de matériel entre chromosomes lors de la formation des gamètes, en particulier au stade prophase de la première division méiotique.
Ce phénomène a été étudié en détail par Barbara McClintock ( prix Nobel en physiologie et médecine en 1983) et s'appelait cross-over. Traverser- ce n'est rien de plus qu'un échange de régions homologues entre chromosomes. Il s'avère que chaque chromosome spécifique, lorsqu'il est transmis de génération en génération, ne reste pas inchangé : il peut « emporter avec lui » une section homologue de son chromosome apparié, lui donnant, à son tour, une section de son ADN.
Dans le cas des humains, il peut être assez difficile d'établir la liaison des gènes, ainsi que d'identifier les croisements en raison de l'impossibilité de croisements arbitraires (vous ne pouvez pas forcer les gens à produire une progéniture conformément à certaines tâches scientifiques !), donc ces données ont été obtenues principalement sur les plantes, les insectes et les animaux. Cependant, grâce à l'étude de familles nombreuses dans lesquelles plusieurs générations sont présentes, des exemples de liaison autosomique (c'est-à-dire de transmission conjointe de gènes situés sur des autosomes) sont connus chez l'homme. Par exemple, il existe un lien étroit entre les gènes qui contrôlent le facteur Rh (Rh) et le système antigénique du groupe sanguin MNS. Chez l'homme, les cas de liaison de certaines caractéristiques avec le sexe, c'est-à-dire en relation avec les chromosomes sexuels, sont mieux connus.
Le franchissement améliore généralement variabilité combinatoire, c’est-à-dire qu’il contribue à une plus grande diversité de génotypes humains. À cet égard, ce processus est d'une grande importance pour. En utilisant le fait que plus les gènes sont éloignés les uns des autres sur le même chromosome, plus ils sont susceptibles de se croiser, Alfred Sturtevant a construit les premières cartes des chromosomes de la drosophile. Aujourd'hui, des cartes physiques complètes de tous les chromosomes humains ont été obtenues, c'est-à-dire que l'on sait dans quel ordre et quels gènes se trouvent sur eux.
Les lois de Mendel
Schéma des première et deuxième lois de Mendel. 1) Une plante à fleurs blanches (deux exemplaires de l'allèle récessif w) est croisée avec une plante à fleurs rouges (deux exemplaires de l'allèle dominant R). 2) Toutes les plantes descendantes ont des fleurs rouges et le même génotype Rw. 3) Lors de l'autofécondation, 3/4 des plantes de la deuxième génération ont des fleurs rouges (génotypes RR + 2Rw) et 1/4 ont des fleurs blanches (ww).
Les lois de Mendel- ce sont les principes de transmission des caractères héréditaires des organismes parents à leurs descendants, issus des expériences de Gregor Mendel. Ces principes ont constitué la base de la génétique classique et ont ensuite été expliqués comme une conséquence des mécanismes moléculaires de l'hérédité. Bien que trois lois soient généralement décrites dans les manuels de langue russe, la « première loi » n’a pas été découverte par Mendel. Parmi les modèles découverts par Mendel, l’« hypothèse de la pureté des gamètes » revêt une importance particulière.
Histoire
Au début du XIXe siècle, J. Goss, expérimentant avec des pois, montra qu'en croisant des plantes avec des pois bleu verdâtre et des pois blanc jaunâtre de la première génération, on obtenait des pois jaune-blanc. Cependant, au cours de la deuxième génération, les traits qui n'apparaissaient pas dans les hybrides de la première génération et appelés plus tard récessifs par Mendel sont réapparus, et les plantes qui les contenaient ne se sont pas divisées lors de l'autopollinisation.
O. Sarge, menant des expériences sur les melons, les a comparés selon des caractéristiques individuelles (pulpe, peau, etc.) et a également établi l'absence de confusion de caractéristiques qui ne disparaissaient pas chez les descendants, mais étaient seulement redistribuées entre eux. C. Nodin, traversée différentes sortes Datura, retrouve une prédominance des signes de Datura Datula tatula au-dessus de Datura stramonium, et cela ne dépendait pas de quelle plante était la mère et laquelle était le père.
Ainsi, au milieu du XIXe siècle, le phénomène de dominance, l'uniformité des hybrides dans la première génération (tous les hybrides de la première génération se ressemblent), le clivage et la combinatoire des caractères dans la deuxième génération sont découverts. Cependant, Mendel, appréciant grandement le travail de ses prédécesseurs, a souligné qu'ils n'avaient pas trouvé de loi universelle pour la formation et le développement des hybrides et que leurs expériences n'étaient pas suffisamment fiables pour déterminer des rapports numériques. La découverte d'une méthode aussi fiable et l'analyse mathématique des résultats, qui ont contribué à créer la théorie de l'hérédité, constituent le principal mérite de Mendel.
Les méthodes de Mendel et l'avancement des travaux
- Mendel a étudié comment les traits individuels sont hérités.
- Mendel a choisi parmi toutes les caractéristiques uniquement les caractéristiques alternatives - celles qui avaient deux options clairement différentes dans ses variétés (les graines sont lisses ou ridées ; il n'y a pas d'options intermédiaires). Un tel rétrécissement conscient du problème de recherche a permis d'établir clairement les schémas généraux d'héritage.
- Mendel a planifié et réalisé une expérience à grande échelle. Il a reçu 34 variétés de pois d'entreprises semencières, parmi lesquelles il a sélectionné 22 variétés « pures » (ne produisant pas de ségrégation selon les caractéristiques étudiées lors de l'autopollinisation). Ensuite, il a procédé à une hybridation artificielle des variétés et a croisé les hybrides résultants entre eux. Il a étudié l'héritage de sept caractères, étudiant un total d'environ 20 000 hybrides de deuxième génération. L'expérience a été facilitée par un choix réussi de l'objet : les pois sont normalement autogames, mais l'hybridation artificielle est facile à réaliser.
- Mendel a été l'un des premiers en biologie à utiliser des méthodes quantitatives précises pour analyser les données. Fort de sa connaissance de la théorie des probabilités, il s'est rendu compte de la nécessité d'analyser un grand nombre de croisements pour éliminer le rôle des écarts aléatoires.
Mendel a qualifié de dominance la manifestation du trait d'un seul des parents chez les hybrides.
Loi d'uniformité des hybrides de première génération(Première loi de Mendel) - lors du croisement de deux organismes homozygotes appartenant à des lignées pures différentes et différant l'un de l'autre par une paire de manifestations alternatives du trait, toute la première génération d'hybrides (F1) sera uniforme et portera la manifestation du trait de caractère de l'un des parents.
Cette loi est également connue sous le nom de « loi de dominance des traits ». Sa formulation est basée sur le concept ligne épurée par rapport à la caractéristique étudiée - sur langue moderne cela signifie que les individus sont homozygotes pour ce trait. Mendel a formulé la pureté d'un caractère comme l'absence de manifestations de caractères opposés chez tous les descendants de plusieurs générations d'un individu donné lors de l'autopollinisation.
En croisant des lignées pures de pois à fleurs violettes et de pois à fleurs blanches, Mendel a remarqué que les descendants des plantes qui ont émergé étaient tous à fleurs violettes, sans aucune blanche parmi eux. Mendel a répété l'expérience plus d'une fois et a utilisé d'autres signes. S’il croisait des pois avec des graines jaunes et vertes, tous les descendants auraient des graines jaunes. S'il croisait des pois avec des graines lisses et ridées, la progéniture aurait des graines lisses. La progéniture des plantes hautes et courtes était grande. Ainsi, les hybrides de première génération sont toujours uniformes dans cette caractéristique et acquièrent la caractéristique de l'un des parents. Ce signe (plus fort, dominant), supprimait toujours l'autre ( récessif).
Codominance et dominance incomplète
Certains caractères opposés ne sont pas dans la relation de dominance complète (quand l'un supprime toujours l'autre chez les individus hétérozygotes), mais dans la relation dominance incomplète. Par exemple, lorsque l'on croise des lignées pures de muflier à fleurs violettes et blanches, les individus de la première génération ont des fleurs roses. Lorsque des lignées pures de poulets andalous noirs et blancs sont croisées, des poulets gris naissent dès la première génération. À dominance incomplète, les hétérozygotes ont des caractéristiques intermédiaires entre celles des homozygotes récessifs et dominants.
Le phénomène dans lequel le croisement d'individus hétérozygotes conduit à la formation de descendants, dont certains portent un trait dominant et d'autres un trait récessif, est appelé ségrégation. Par conséquent, la ségrégation est la répartition des traits dominants et récessifs parmi la progéniture dans un certain rapport numérique. Le caractère récessif ne disparaît pas chez les hybrides de première génération, mais est seulement supprimé et apparaît dans la deuxième génération hybride.
Explication
Loi de la pureté des gamètes: chaque gamète ne contient qu'un seul allèle issu d'une paire d'allèles d'un gène donné de l'individu parent.
Normalement, le gamète est toujours pur du deuxième gène de la paire allélique. Ce fait, qui n'a pas pu être fermement établi à l'époque de Mendel, est également appelé hypothèse de pureté des gamètes. Cette hypothèse a ensuite été confirmée par des observations cytologiques. De toutes les lois successorales établies par Mendel, cette « Loi » est la plus générale par nature (elle s'accomplit dans le plus large éventail de conditions).
Loi de l'héritage indépendant des caractéristiques
Illustration de l'héritage indépendant des traits
Définition
Droit de l'héritage indépendant(Troisième loi de Mendel) - lors du croisement de deux individus homozygotes qui diffèrent l'un de l'autre par deux (ou plus) paires de traits alternatifs, les gènes et leurs traits correspondants sont hérités indépendamment les uns des autres et sont combinés dans toutes les combinaisons possibles (comme dans le croisement monohybride ). Lorsqu'on croisait des plantes différant par plusieurs caractères, comme les fleurs blanches et violettes et les pois jaunes ou verts, l'hérédité de chaque caractère suivait les deux premières lois et chez la progéniture, elles étaient combinées de telle manière que si leur hérédité se produisait indépendamment de l'un l'autre. La première génération après croisement avait un phénotype dominant pour tous les caractères. Dans la deuxième génération, une division des phénotypes a été observée selon la formule 9:3:3:1, c'est-à-dire que 9:16 étaient à fleurs violettes et pois jaunes, 3:16 étaient à fleurs blanches et pois jaunes, 3 : 16 étaient à fleurs violettes et pois verts, 1 :16 à fleurs blanches et pois verts.
Explication
Mendel a découvert des caractères dont les gènes étaient situés dans différentes paires de chromosomes homologues du pois. Au cours de la méiose, des chromosomes homologues de différentes paires sont combinés au hasard dans les gamètes. Si le chromosome paternel de la première paire pénètre dans le gamète, alors avec une probabilité égale, les chromosomes paternels et maternels de la deuxième paire peuvent pénétrer dans ce gamète. Par conséquent, les traits dont les gènes sont situés dans différentes paires de chromosomes homologues sont combinés indépendamment les uns des autres. (Il s'est avéré plus tard que sur les sept paires de caractères étudiées par Mendel chez le pois, qui possède un nombre diploïde de chromosomes 2n=14, les gènes responsables d'une des paires de caractères étaient situés sur le même chromosome. Cependant, Mendel n'a pas découvert de violation de la loi de l'hérédité indépendante, puisque le lien entre ces gènes n'a pas été observé en raison de la grande distance qui les sépare).
Dispositions fondamentales de la théorie de Mendel sur l'hérédité
Dans l'interprétation moderne, ces dispositions sont les suivantes :
- Facteurs héréditaires discrets (séparés, non mélangeables) - les gènes sont responsables des traits héréditaires (le terme « gène » a été proposé en 1909 par V. Johannsen)
- Chaque organisme diploïde contient une paire d'allèles d'un gène donné responsable d'un trait donné ; l'un d'eux est reçu du père, l'autre de la mère.
- Les facteurs héréditaires sont transmis aux descendants par les cellules germinales. Lorsque les gamètes se forment, chacun d’eux ne contient qu’un seul allèle de chaque paire (les gamètes sont « purs » dans le sens où ils ne contiennent pas le deuxième allèle).
Conditions pour l'accomplissement des lois de Mendel
Selon les lois de Mendel, seuls les traits monogéniques sont hérités. Si plusieurs gènes sont responsables d’un trait phénotypique (et de la majorité absolue de ces traits), son mode de transmission est plus complexe.
Conditions pour respecter la loi de ségrégation lors de la traversée monohybride
La répartition 3:1 par phénotype et 1:2:1 par génotype est réalisée approximativement et uniquement dans les conditions suivantes.
























