Монооксигеназные реакции необходимы для:
1. специфических превращений аминокислот, например, для синтеза тирозина из фенилаланина (фермент – фенилаланингидроксилаза);
2. синтеза холестерола, желчных кислот в печени; стероидных гормонов в коре надпочечников, яичниках, плаценте, семенниках; витамина D 3 в почках;
3. обезвреживания чужеродных веществ (ксенобиотиков) в печени.
Ферменты монооксигеназного пути окисления локализованы в мембранах эндоплазматического ретикулума (при гомогенизации тканей эти мембраны превращаются в микросомы – мембранные пузырьки). Поэтому монооксигеназный путь окисления называют микросомальным окислением.
Микросомальное окисление представляет короткую электронтранспортную цепь, включающую НАДФ, ФАД, ФМН, цитохром Р 450 .
Микросомальная система включает два фермента: цитохром Р 450 и НАДФН-цитохром-Р 450 -редуктазу.
НАДФН-цитохром Р 450 – редуктаза – флавопротеин, в качестве простетической группы содержит два кофермента ФАД и ФМН.
Цитохром Р 450 – гемопротеин, содержит простетическую группу гем и участки связывания для кислорода и субстрата. Восстановленный цитохром Р 450 имеет максимум поглощения при 450 нм. Выполняет две функции: связывание окисляемого субстрата и активация молекулярного кислорода.

Рис. 11.1. Схема микросомального окисления
Микросомальное окисление протекает в несколько этапов:
1. связывание в активном центре цитохрома Р 450 субстрата RН;
2. присоединение первого электрона и восстановление железа в геме до Fe 2+ ; изменение валентности железа увеличивает сродство комплекса Р 450 – Fe 2+ RH к молекуле кислорода; присоединение второго электрона к молекуле кислорода и образование неустойчивого пероксикомплекса Р 450 –Fe 2+ О 2 - RH;
3. Fe 2+ окисляется, при этом электрон присоединяется к молекуле кислорода; восстановленный атом кислорода (О 2 -) связывает два протона (донор протонов – НАДФН+Н +) и образуется 1 молекула воды; второй атом кислорода участвует в гидроксилировании субстрата RH; гидроксилированный субстрат ROH отделяется от фермента.
В результате гидроксилирования гидрофобный субстрат становится более полярным, повышается его растворимость и возможность выведения из организма с мочой. Так окисляются многие ксенобиотики, лекарственные вещества.
В редких случаях в результате гидроксилирования токсичность соединения увеличивается. Например, при окислении нетоксичного бензпирена (содержится в табачном дыму, копченостях) образуется токсичный оксибензпирен, который является сильным концерогеном, индуцирующим злокачественное перерождение клеток.
В митохондриях содержится монооксигеназная система, которая выполняет биосинтетическую функцию: синтез холестерола; стероидных гормонов (кора надпочечников, яичники, плацента, семенники); желчных кислот (печень); образование витамина D 3 (почки).
Рис. 12. Схема переноса электронов хлорофилла в процессе синтеза АТФ (фосфорилирования). При нециклическом фосфорилировании за счет энергии ФС II идет синтез АТФ, а за счет ФС I идет синтез НАДФ · 2Н. При циклическом фосфорилировании работает только ФС I и происходит синтез АТФ
Под действием света происходит возбуждение молекул хлорофилла ФС I и ФС II, которые, поглощая кванты света, испускают возбужденные электроны. За счет энергии электронов, летящих по цепи ферментов от ФС II к ФС I, происходит синтез молекул АТФ. Этот процесс называется фосфорилированием и происходит в присутствии фермента АТФ-синтетазы.
АДФ + Фн АТФ
где Фн - неорганический фосфат H3 PO4 .
Электроны ФС II, потерявшие энергию, попадают на ФС I, которая является их конечным акцептором.
Электроны, покидающие хлорофилл фотосистемы ФС I при поглощении света, также попадают на электроннотранспортную цепь. Но за счет их энергии идет соединение протонов Н+ с переносчиком НАДФ+ .
Электроны, которые теряет ФС I, восполняются электронами ФС II, а электроны, отданные молекулой воды при фотолизе, восполняют электроны, потерянные хлорофиллом ФС II, и он также восстанавливается:
хлорофилл+2 + 2ē хлорофилл.
На мембране имеются специальные протонные каналы, по которым в определенный момент ионы водорода могут переходить из Н+ -резервуара тилакоида в строму хлоропласта. Каналы связаны с ферментом АТФ-синтетазой. Когда возбужденные электроны ФС I, двигаясь по мембране, достигают протонного канала, он открывается и в него устремляются ионы водорода. Этот процесс сопряжен с синтезом АТФ и происходит синхронно.
С наружной стороны мембраны тилакоида, т. е. в строме хлоропласта, скапливаются молекулы переносчика водорода НАДФ+ в окисленном состоянии. Они принимают электроны от ФС I, за счет чего происходит их соединение с ионами водорода Н+ и образование НАДФ · 2H:
НАДФ+ + 2H+ + 2ē НАДФ · 2Н. (4)
Синтез АТФ и НАДФ · 2Н протекает на мембранах тилакоидов и сопряжен с переносом возбужденных электронов по электронно-транспортной цепи. Таким образом, энергия солнца преобразуется в энергию возбужденных электронов, а далее запасается в процессе синтеза в молекулах АТФ и НАДФ · 2Н.
Суммарное уравнение реакций световой фазы:
H2 O + НАДФ+ + 2АДФ + 2Фн НАДФ · 2Н + 2АТФ + 1 /2 O2 .
Темновая фаза
Реакции темновой фазы (рис. 13) протекают в строме хлоропластов, куда поступают молекулы НАДФ · 2Н и АТФ, синтезированные в световую фазу, и углекислый газ из атмосферы. Последовательность циклических реакций этой фазы была описана американским ученым-биохимиком Мэлвином Кальвином и получила название цикла Кальвина. Здесь происходит связывание молекул CO2 , активирование соединений за счет АТФ (фосфорилирование), восстановление углерода водородом из НАДФ · 2Н и синтез глюкозы. Источником энергии являются синтезированные на первой стадии молекулы АТФ.
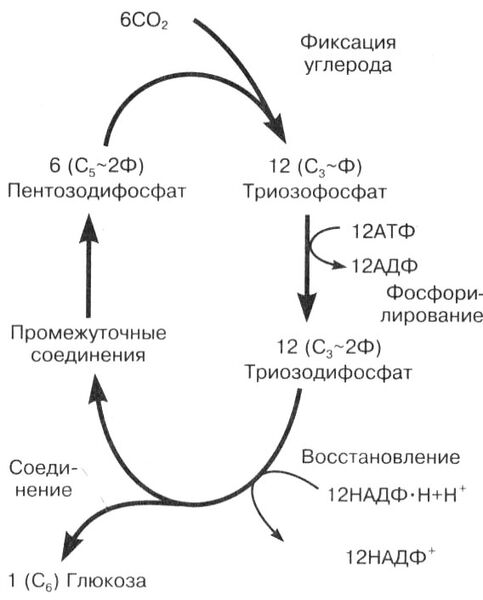
Рис. 13. Общая схема темновых реакций фотосинтеза. Цикл Кальвина
В строме хлоропласта постоянно присутствует пятиуглеродный углевод (пентоза), связанный с двумя остатками фосфорной кислоты - рибулозодифосфат.
Образующееся шестиуглеродное соединение неустойчиво и сразу же распадается на два триозофосфата.
С5 -углевод-2Ф + CO2 С6 -углевод-2Ф 2С3 -Ф
2С3 Ф + 2АТФ 2АДФ + 2С3 ~2Ф.

Рис. 14. Фиксация углерода, его фосфорилирование и восстановление
После этого происходит восстановление триозодифосфатов молекулами НАДФ · 2Н:
2С3 ~2Ф + 2НАДФ · 2Н 2С3 + 2НАДФ+ + 2Ф.
Две молекулы триозы соединяются между собой, и образуется глюкоза, которая может в дальнейшем превращаться в сахарозу, крахмал и другие полисахариды:
2С3 С6 Н12 O6 .
Часть молекул триоз может использоваться для синтеза аминокислот, глицерина, высших жирных кислот.
Частично триозы продолжают участвовать в циклических реакциях и превращаются вновь в пентозу, которая замыкает цикл.
В реакции участвуют одновременно шесть молекул каждого вещества. Таким образом, для синтеза одной полностью новой молекулы глюкозы цикл должен повториться шесть раз, т. е. должно усвоиться шесть молекул CO2 .
Освобожденные молекулы АТФ и НАДФ+ вновь возвращаются к мембранам тилакоидов для участия в световых реакциях.
Влияние на скорость фотосинтеза различных факторов
Интенсивность фотосинтеза зависит от целого ряда факторов. Во-первых, это длина световой волны.
Наиболее интенсивен процесс в ультрафиолетовой и красной части спектра. Кроме того, скорость фотосинтеза зависит от степени освещенности, и до определенной величины она возрастает пропорционально освещенности, но далее уже не зависит от нее (рис. 15).

Рис. 15. Влияние различных факторов на скорость фотосинтеза
Другим фактором является концентрация углекислого газа. Чем выше количество углекислого газа, тем интенсивнее идет процесс фотосинтеза. В обычных условиях недостаток углекислого газа является главным лимитирующим фактором, так как в атмосферном воздухе содержится небольшой его процент. Однако в условиях теплиц можно устранить этот дефицит, что благоприятно влияет на интенсивность роста и развития растений.
Немаловажным фактором является температура.
Все реакции фотосинтеза катализируются ферментами, для которых оптимальной температурой является интервал 25–30 °C. При низких температурах скорость действия ферментов резко снижается.
Вода является также важным фактором, влияющим на фотосинтез. Однако оценить количественно этот фактор невозможно, так как она участвует во многих других обменных процессах.
Для темновых реакций, которые протекают в строме, свет не нужен. Восстановление СО 2 происходит за счет энергии (АТФ) и восстановительной силы (НАДФ·Н 2), образующихся при световых реакциях. Темновые реакции контролируются ферментами. Последовательность этих реакций была определена в США Кальвином, Бенсоном и Бэссемом в период с 1946 по 1953 г.; в 1961 г. Кальвину была присуждена за эту работу Нобелевская премия.
Эксперименты Кальвина
Работы Кальвина были основаны на использовании радиоактивного изотопа углерода 14 С (период полураспада 5570 лет, см. Приложение 1.3), который стал доступным для исследователей только в 1945 г. Кроме того, Кальвин применял бумажную хроматографию, которая в то время была сравнительно новым, еще мало распространенным методом. Культуры одноклеточной зеленой водоросли хлореллы (Chlorella) выращивали в специальном аппарате (рис. 9.17). Культуру выдерживали с 14 СО 2 в течение различных промежутков времени, затем клетки быстро фиксировали, выливая суспензию в горячий метанол. Растворимые продукты фотосинтеза экстрагировали, концентрировали и разделяли при помощи двумерной хроматографии на бумаге (рис. 9.18 и Приложение 1.8.2). Цель состояла в том, чтобы проследить путь, по которому меченый углерод попадает (через ряд промежуточных продуктов) в конечные продукты фотосинтеза. Положение радиоактивных соединений на бумаге определяли с помощью радиоавтографии : для этого на хроматограмму накладывали фотопленку, чувствительную к излучению 14 С, и она засвечивалась, т. е. чернела, в тех местах, где находились радиоактивные вещества (рис. 9.18). Уже за одну минуту инкубации с 14 СО 2 синтезировались многие сахара и органические кислоты, в том числе различные аминокислоты. Однако Кальвину удалось, используя очень короткие экспозиции - в течение 5 и менее секунд - идентифицировать первый продукт фотосинтеза и установить, что это кислота, содержащая три атома углерода, а именно фосфоглицериновая кислота (ФГК). Затем он выяснил всю цепь промежуточных соединений, по которой передается фиксированный углерод; эти стадии будут рассмотрены позднее. С тех пор эти реакции называют циклом Кальвина (или циклом Кальвина - Бенсона- Бэссема).

Рис. 9.18. А. Фиксация 14 СО 2 У водорослей при кратковременном освещении. Определение продуктов фиксации с помощью хроматографии на бумаге и радиоавтографии. Б. Радиоавтографы продуктов фотосинтеза, получающихся после кратковременного освещения водорослей в присутствии 14 СО 2
9.18. Какие преимущества дает применение долгоживущих радиоактивных изотопов в биологических исследованиях?
9.19. Какие преимущества можно получить, взяв хлореллу вместо высшего растения?
9.20. Почему сосуд аппарата Кальвина имеет плоскую, а не шарообразную форму?
Этапы пути углерода
Фиксация двуокиси углерода:

Акцептором СО 2 служит пятиуглеродный сахар (пентоза) рибулозобисфосфат (т. е. рибулоза с двумя фосфатными группами; раньше это соединение называли рибулозодифосфатом). Присоединение СО 2 к тому или иному веществу называется карбоксилированием , а фермент, катализирующий такую реакцию, - карбоксилазой . Образующийся шестиуглеродный продукт неустойчив и сразу же распадается на две молекулы фосфоглицериновой кислоты (ФГК), которая и является первым продуктом фотосинтеза. Фермент рибулозобисфосфат-карбоксилаза содержится в строме хлоропластов в большом количестве - это фактически самый распространенный в мире белок.
Восстановительная фаза :

ФГК содержит три атома углерода и имеет кислотную карбоксильную группу (-СООН). ТФ - это триозофосфат, или глицеральдегидфосфат (трехуглеродный сахар); он имеет альдегидную группу (-СНО).
Для удаления кислорода из ФГК (т. е. для ее восстановления) используются восстановительная сила НАДФ·Н 2 и энергия АТФ. Реакция протекает в два этапа: сначала расходуется часть АТФ, образовавшегося в ходе световых реакций, а затем используется весь НАДФ·Н 2 , также полученный на свету. Суммарный результат - восстановление карбоксильной группы кислоты (-СООН) до альдегидной группы (-СНО). Продукт реакции - триозофосфат, т. е. трехуглеродный сахар с присоединенной к нему фосфатной группой. В этом соединении больше химической энергии, чем в ФГК, и это первый углевод, который образуется при фотосинтезе.
Регенерация акцептора для СО 2 - рибулозобисфосфата . Часть триозофосфата (ТФ) должна израсходоваться на регенерацию рибулозобисфосфата, который используется в первой реакции. Этот процесс представляет собой сложный цикл, в котором участвуют сахарофосфаты с 3, 4, 5, 6, 7 атомами углерода. Именно здесь и расходуется остальной АТФ. Все темновые реакции суммированы на рис. 9.19. На этом рисунке цикл Кальвина изображен в виде "черного ящика", в который с одной стороны поступают СО 2 и Н 2 О, а с другой стороны выходит триозофосфат. Как видно из этой схемы, остаток АТФ используется для фосфорилирования рибулозобисфосфата, однако детали этой сложной цепи реакций не показаны.

Из рис. 9.19 можно вывести такое суммарное уравнение:

Здесь важно обратить внимание на то, что на образование двух молекул триозофосфата идет шесть молекул СО 2 . Уравнение можно упростить, поделив все коэффициенты на 6:

9.21. Перерисуйте рис. 9.19, указав только число атомов углерода, участвующих в реакциях; например, вместо 6 РиБФ напишите "6 × 5С" и т. д.
Основные сведения о процессе фотосинтеза резюмированы в табл. 9.6.
| Световые реакции | Темновые реакции | |
| Локализация в хлоропластах | Тилакоиды | Строма |
| Реакции | Фотохимические, т. е. требуют света. Световая энергия вызывает перенос электронов от "доноров" электронов к их "акцепторам" либо по нециклическому, либо по циклическому пути. Участвуют две фотосистемы - Ι и ΙΙ. В них находятся молекулы хлорофиллов, которые при поглощении энергии света испускают электроны. Вода служит донором электронов для нециклического пути. Перенос электронов приводит к образованию АТФ (фотофосфорилированию) и НАДФ·Н 2 (см. также табл. 9.5). | Не требуют света. СО 2 фиксируется, когда связывается с пятиуглеродным акцептором - рибулозобисфосфатом (РиБФ); при этом образуются две молекулы трехуглеродного соединения-фосфоглицериновой кислоты (ФГК), первого продукта фотосинтеза. Происходит целый ряд реакций, в совокупности называемых циклом Кальвина; при этом регенерируется акцептор для СО 2 -РиБФ, а ФГК восстанавливается, превращаясь в сахар (см. также рис. 9.19). |
| Комбинированные уравнения |
Как понятно из названия, фотосинтез по своей сути являет собой природный синтез органических веществ, превращая СО2 из атмосферы и воду в глюкозу и свободный кислород.
При этом необходимо наличие энергии солнечного света.
Химическое уравнение процесса фотосинтеза в общем можно представить в следующем виде:
Фотосинтез имеет две фазы: темную и световую. Химические реакции темной фазы фотосинтеза существенно отличаются от реакций световой фазы, однако темная и световая фаза фотосинтеза зависят друг от друга.
Световая фаза может происходить в листьях растений исключительно при солнечном свете. Для темной же необходимо наличие углекислого газа, именно поэтому растение все время должно поглощать его из атмосферы. Все сравнительные характеристики темной и световой фаз фотосинтеза будут предоставлены ниже. Для этого была создана сравнительная таблица «Фазы фотосинтеза».
Световая фаза фотосинтеза
Основные процессы в световой фазе фотосинтеза происходят в мембранах тилакоидов. В ней участвуют хлорофилл, белки-переносчики электронов, АТФ-синтетаза (фермент, ускоряющий реацию) и солнечный свет.
Далее механизм реакции можно описать так: когда солнечный свет попадает на зеленые листья растений, в их структуре возбуждаются электроны хлорофилла (заряд отрицательный), которые перейдя в активное состояние, покидают молекулу пигмента и оказываются на внешней стороне тилакоида, мембрана которого заряжена также отрицательно. В то же время молекулы хлорофилла окисляются и уже окисленные они восстанавливаются, отбирая таким образом электроны у воды, которая находится в структуре листа.
Этот процесс приводит к тому, что молекулы воды распадаются, а созданные в результате фотолиза воды ионы, отдают свои электроны и превращаются в такие радикалы ОН, которые способны проводить дальнейшие реакции. Далее эти реакционноспособные радикалы ОН объединяются, создавая полноценные молекулы воды и кислород. При этом свободный кислород выходит во внешнюю среду.
В результате всех этих реакций и превращений, мембрана тилакоида листа с одной стороны заряжается положительно (за счет иона Н+), а с другой — отрицательно (за счет электронов). Когда разность между этими зарядами в двух сторонах мембраны достигает больше 200 мВ, протоны проходят через специальные каналы фермента АТФ-синтетазы и за счет этого происходит превращение АДФ до АТФ (в результате процесса фосфорилизации). А атомный водород, который освобождается из воды, восстанавливает специфический переносчик НАДФ+ до НАДФ·Н2. Как видим, в результате световой фазы фотосинтеза происходит три основных процесса:
- синтез АТФ;
- создание НАДФ·Н2;
- образование свободного кислорода.
Последний освобождается в атмосферу, а НАДФ·Н2 и АТФ берут участие в темной фазе фотосинтеза.
Темная фаза фотосинтеза
Темная и световая фазы фотосинтеза характеризуются большими затратами энергии со стороны растения, однако темная фаза протекает быстрее и требует меньше энергии. Для реакций темной фазы не нужен солнечный свет, поэтому они могут происходить и днем и ночью.
Все основные процессы этой фазы протекают в строме хлоропласта растения и являют собой своеобразную цепочку последовательных превращений углекислого газа из атмосферы. Первая реакция в такой цепи – фиксация углекислого газа. Чтобы она проходила более плавно и быстрее, природой был предусмотрен фермент РиБФ-карбоксилаза, который катализирует фиксацию СО2.
Далее происходит целый цикл реакций, завершением которого является преобразование фосфоглицериновой кислоты в глюкозу (природный сахар). Все эти реакции используют энергию АТФ и НАДФ Н2, которые были созданы в световой фазе фотосинтеза. Помимо глюкозы в результате фотосинтеза образуются также и другие вещества. Среди них разные аминокислоты, жирные кислоты, глицерин, а также нуклеотиды.
Фазы фотосинтеза: таблица сравнений
| Критерии сравнения | Световая фаза | Темная фаза |
| Солнечный свет | Обязателен | Необязателен |
| Место протекание реакций | Граны хлоропласта | Строма хлоропласта |
| Зависимость от источника энергии | Зависит от солнечного света | Зависит от АТФ и НАДФ Н2, образованных в световой фазе и от количества СО2 из атмосферы |
| Исходные вещества | Хлорофилл, белки-переносчики электронов, АТФ-синтетаза | Углекислый газ |
| Суть фазы и что образуется | Выделяется свободный О2, образуется АТФ и НАДФ Н2 | Образование природного сахара (глюкозы) и поглощение СО2 из атмосферы |
Фотосинтез — видео
Процесс фотосинтеза состоит из двух последовательных фаз: световой и темновой.
Световая фаза. Расположенные в хлоропластах молекулы хлорофилла поглощают лучи определенного участка спектра (красные и фиолетовые). Поглотив квант света, молекула хлорофилла возбуждается. Квант света срывает электрон с ее орбиты, в результате чего молекула хлорофилла окисляется, а электрон присоединяется к молекуле переносчика электронов .
В процессе последующих окислительно-восстановительных реакций электрон транспортируется другими переносчиками с более низкими окислительно-восстановительными потенциалами. Освобожденная при этом энергия используется прежде всего на образование АТФ из АДФ. Фотосинтетическое фосфорилирование – присоединение неорганического фосфата к АДФ с использованием энергии света . Различают циклическое и нециклическое фосфорилирование. При циклическом фосфорилировании восстановление молеул хлорофилла проходит за счет возвращения его собственных электронов, предварительно сорванных с орбиты квантом света. В этом случае образуется только АТФ за счет энергии электронов активированного светом хлорофилла.

В результате нециклического фосфорилирования хлорофилл восстанавливается за счет электронов гидроксильных ионов воды, которая предварительно подвергается фотолизу – фотохимическому расщеплению на ион водорода и гидроксил. На этом пути энергия электрона используется для «зарядки» АТФ, а протоны воды, соединяясь с НАДФ, образуют его восстановленную формулу – НАДФ. Н (с участием электронов, оторванных от молекулы хлорофилла). Кроме такого восстановления образуется и кислород из групп ОН. (рис).
Кроме АТФ энергия аккумулируется и в системе НАДФ – НАДФ. Н. Химическая энергия, аккумулированная в АТФ и НАДФ. Н используется в дальнейшем для синтеза органических соединений.
Процесс движения электронов при фосфорилировании не сопровождается мгновенной отдачей энергии. Это объясняется тем, что электроны с высших энергетических орбит могут переноситься от молекулы к молекуле системой переносчиков электронов, которыми являются пластохиноны, цитохромы, ферредоксин и другие соединения. Такая миграция электронов замедляет отдачу ими энергии, что биологически более целесообразно, чем одномоментная отдача энергии, которую соответствующие системы не успели бы «переработать».
Световая фаза проходит в мембране гран (рис.2).Под действием кванта света хлорофилл теряет электрон и переходит в возбужденное состояние:
Хл свет Хл * + е - .
Эти электроны передаются переносчиками на наружную (обращенную к матриксу) поверхность мембраны тилакоидов, где накапливаются. Одновременно внутри полостей тилакоидов происходит фотолиз:
Н 2 О свет Н + + ОН - .
Ионы гидроксила отдают свои электроны, превращаясь в реакционноспособные радикалы ОН:
ОН - – е - = ОН.

Образующиеся электроны передаются переносчиками к молекулам хлорофилла и восстанавливают их, а радикалы ОН объединяются, образуя перекись водорода, которая на свету быстро разлагается на воду и свободный кислород:
4ОН = 2Н 2 О 2 ; 2Н 2 О 2 = 2Н 2 О + О 2 .
Протоны водорода, образовавшиеся при фотолизе воды, не могут проникнуть через мембрану граны и накапливаются внутри нее, создавая и пополняя протонный резервуар . В результате внутренняя поверхность мембраны граны заряжается положительно (за счет Н +), а наружная – отрицательно (за счет е -). По мере накопления по обе стороны мембраны противоположно заряженных частиц нарастает разность потенциалов. При достижении ее критической величины сила электростатического поля начинает проталкивать протоны через канал АТФ-синтетазы, расположенный в АТФсоме. На выходе из протонного канала создается высокий уровень энергии, которая используется для фосфорилирования имеющихся в матриксе молекул АДФ:
АДФ + Ф = АТФ.

Ионы водорода, оказавшись на наружной поверхности мембраны тилакоида, встречаются там с электронами, образуя атомарный водород, который идет на восстановление специфического переносчика НАДФ + (никотинамидадениндинуклеотидфосфата ):
2Н + + 4е - + НАДФ + = НАДФ. Н 2 .
Таким образом цикл световых реакций фотосинтеза представляет собой индуцированное светом перенесение электронов в реакциях циклического и нециклического фосфорилирования, во время которых происходят три процесса: образование кислорода вследствие разложения воды, синтез АТФ и образование атомов водорода в форме НАДФ. Н 2 . Кислород диффундирует в атмосферу, а 3 АТФ и 2 НАДФ. Н 2 транспортируются в матрикс пластид и участвуют в процессах темновой фазы.
Поглощенная световая энергия может переводить электроны молекул хлорофилла только на высший уровень. Если такая система не имеет компонентов электронно-транспортной цепи, то электроны за очень короткое время возвращаются к предыдущему состоянию и молекулы отдают освобожденную энергию в виде света флуоресценции.
Темновая (термохимическая) фаза фотосинтеза протекает в матриксе хлоропласта как на свету, так и в темноте, и представляет собой ряд последовательных преобразований СО 2 , поступающего из воздуха. НАДФ. Н 2 и АТФ, образованные во время световой фазы, запускает циклический механизм темновых реакций – цикл Кальвина.
Углекислый газ присоединяется к акцептору – рибулозо-1,5-дифосфату . Образуется нестойкое шестиуглеродное (С6) соединение, которое распадается на две молекулы фосфоглицериновой кислоты (С3): начинается так называемый углеродный цикл фотосинтеза , который включает 13 и более этапов – последовательных и взаимообусловленных реакций, в результате которых образуются богатые энергией углеводы, а немного позже – жиры и белки. Важнейшие и узловые реакции и этапы этих преобразований:
- фосфоглицериновая кислота , используя кислотный остаток и энергию ранее синтезированной АТФ, превращается в дифосфоглицериновую кислоту;
- дифосфоглицериновая кислота с помощью водорода от НАДФ. Н 2 восстанавливается до фосфоглицеринового альдегида; последний может изомеризироваться в фосфодиоксиацетон, который может быть исходным для образования глицерола и жирных кислот;
Остатки фосфоглицеринового альдегида соединяются с образованным фосфодиоксиацетоном , образуя фруктозодифосфат – исходное вещество для синтеза растворимых углеводов, крахмала и других полисахаридов;
От части молекул фруктозодифосфата отщепляется по одному остатку фосфорной кислоты, вследствие чего образуется фруктозомонофосфат (фруктозо-6-фосфат);
- фруктозо-6-фосфат соединяется с фосфоглицериновым альдегидом, образуя одну молекулу эритрозофосфата и одну молекулу ксилулозофосфата; эти фосфорилированные 4- и 5-углеродные сахара в свою очередь в результате сложных реакций превращаются в некоторые аминокислоты (триптофан) и далее в НАДФ и азотистые основания; образование других аминокислот начинается на этапе образования фосфоглицериновой кислоты, часть которой отделяется;
- эритрозофосфат проходит ряд реакций, в результате которых образуется рибозофосфат;
Рибозофосфат с участием АТФ фосфорилируется и превращается в рибулезо-1,5-дифосфат и начинается новый цикл.
В результате шести циклов темновых реакций синтезируется одна молекула глюкозы и другие важные соединения, для чего необходимо 18 молекул АТФ и 12 молекул НАДФ. Н 2 , т. е. на восстановление одной молекулы СО 2 необходимо 3 АТФ и 2 НАДФ. Н 2 .
| Фотосинтез | Стадии фотосинтеза | |
| Световая стадия (или энергетическая) | Темновая стадия (или метаболическая) | |
| Место протекание реакции | В квантосомах мембран тилактоидов, протекает на свету. | Осуществляется вне тилактоидов, в водной среде стромы. |
| Начальные продукты | Энергия света, вода (Н2О), АДФ, хлорофилл | СО2, рибулозодифосфат, АТФ, НАДФН2 |
| Суть процесса | Фотолиз воды, фосфорилирование В световой стадии фотосинтеза энергия света трансформируется в химическую энергию АТФ, а бедные энергией электроны воды переходят в богатые энергией электроны НАДФ·Н2. Побочным веществом, образующимся в ходе световой стадии, является кислород. Реакции световой стадии получили название "световых реакций". | Карбоксилирование, гидрирование, дефосфорилирование В темновой стадии фотосинтеза протекают "темновые реакции" при которых наблюдается восстановительный синтез глюкозы из CO2. Без энергии световой стадии темновая стадия невозможна. |
| Конечные продукты | О2, АТФ, НАДФН2 Богатые энергией продукты световой реакции - АТФ и НАДФ·Н2 далее используются в темновой стадии фотосинтеза. | С6Н12О6 |
Взаимосвязь между световой и темновой стадиями можно выразить схемой
 Процесс фотосинтеза эндергонический, т.е. сопровождается увеличением свободной энергии, поэтому требует значительного количества энергии, подведенной извне. Суммарное уравнение фотосинтеза:
6СО2 + 12Н2О--->С6Н12О62 + 6Н2О + 6О2 + 2861 кДж/моль.
Процесс фотосинтеза эндергонический, т.е. сопровождается увеличением свободной энергии, поэтому требует значительного количества энергии, подведенной извне. Суммарное уравнение фотосинтеза:
6СО2 + 12Н2О--->С6Н12О62 + 6Н2О + 6О2 + 2861 кДж/моль.
|
Значение фотосинтеза :
1. «Консервирование» солнечной энергии : в процессе фотосинтеза световая энергия трансформируется в энергию химических связей синтезированных органических соединений. Такая форма энергии сохраняется до момента распада органических соединений, т. е. неопределенно долго. При полном окислении 1 г глюкозы выделяется 669 ккал, т. е. столько, сколько поглощается при ее образовании, Тепловая энергия, выделяющаяся при сгорании нефти, угля, торфа, древесины, - все это энергия Солнца, усвоенная и преобразованная растениями.
2. Образование свободного кислорода : свободный кислород необходим для дыхания всех аэробов – один человек за сутки потребляет 500 л кислорода, а за год – более 180 000 л.; кислородное дыхание обеспечивает по сравнению с анаэробным более высокий жизненный уровень, быстрый рост, интенсивное размножение, широкое расселение вида, т. е. биологический прогресс.
3. Образование разнообразных органических соединений : растения синтезируют углеводы, белки, жиры, которые служат пищей для животных и человека, сырьем для промышленности; растения образуют каучук, гуттаперчу, эфирные масла, смолы, дубильные вещества, алколоиды и т. п.; продукты переработки растительного сырья – это ткани, бумага, красители, лекарственные и взрывчатые вещества, искусственное волокно, строительные материалы и др.
4. Извлечение из атмосферы углекислого газа : ежегодно растения поглощают 15,6х10 10 тонн углекислого газа (1/16 часть мирового запаса) и 220 млрд. тонн воды. Количество органического вещества на Земле составляет 10 14 тонн, причем масса растений относится к массе животных как 2200:1.
























